IGMIN: We're glad you're here. Please click 'create a new query' if you are a new visitor to our website and need further information from us.
If you are already a member of our network and need to keep track of any developments regarding a question you have already submitted, click 'take me to my Query.'
Welcome to IgMin Research – an Open Access journal uniting Biology, Medicine, and Engineering. We’re dedicated to advancing global knowledge and fostering collaboration across scientific fields.
At IgMin Research, we bridge the frontiers of Biology, Medicine, and Engineering to foster interdisciplinary innovation. Our expanded scope now embraces a wide spectrum of scientific disciplines, empowering global researchers to explore, contribute, and collaborate through open access.
Welcome to IgMin, a leading platform dedicated to enhancing knowledge dissemination and professional growth across multiple fields of science, technology, and the humanities. We believe in the power of open access, collaboration, and innovation. Our goal is to provide individuals and organizations with the tools they need to succeed in the global knowledge economy.
IgMin Publications Inc., Suite 102, West Hartford, CT - 06110, USA
Medicine GroupLiterature ReviewArticle ID: igmin310
Engineered Escherichia coli Strains as Therapeutic Agents in Reactive Oxygen Species (ROS)-Mediated Glioblastoma Treatment: A Systematic Review of Mechanisms, Efficacy and Challenges
Glioblastoma multiforme (GBM) remains one of the most aggressive and treatment-resistant brain tumors, characterized by a hostile microenvironment and poor prognosis. Recent advances in synthetic biology have led to the engineering of Escherichia coli (E. coli) strains as living therapeutics capable of targeting GBM via Reactive Oxygen Species (ROS)-mediated mechanisms. This systematic review evaluates the mechanisms by which engineered E. coli modulate ROS, their therapeutic efficacy in preclinical models, and challenges in delivery, safety, and regulation. Engineered strains show promise through prodrug conversion, direct ROS production, and immune activation, especially when combined with nanocarriers and immune checkpoint inhibitors. However, tumor heterogeneity, biosafety concerns, and regulatory complexities remain significant hurdles. Advancing toward clinical translation will require robust biocontainment systems, adaptable genetic circuits, and validation in humanized preclinical models.
Glioblastoma Multiforme (GBM) is the most aggressive and lethal primary brain tumor in adults, characterized by rapid proliferation, extensive infiltration into surrounding brain tissue, marked genetic heterogeneity, and resistance to conventional therapies [11Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.]. Notwithstanding the existing standard of treatment, comprising maximum surgical resection, subsequent concomitant radiation, and temozolomide chemotherapy, median survival remains bleak, often fluctuating between 12 and 15 months [22Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, Gorlia T, Allgeier A, Lacombe D, Cairncross JG, Eisenhauer E, Mirimanoff RO; European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups; National Cancer Institute of Canada Clinical Trials Group. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005 Mar 10;352(10):987-96. doi: 10.1056/NEJMoa043330. PMID: 15758009.,33Ostrom QT, Price M, Neff C, Cioffi G, Waite KA, Kruchko C, Barnholtz-Sloan JS. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2015-2019. Neuro Oncol. 2022 Oct 5;24(Suppl 5):v1-v95. doi: 10.1093/neuonc/noac202. PMID: 36196752; PMCID: PMC9533228.]. This poor prognosis is largely attributed to the tumor’s capacity to evade immune surveillance, its inherent radioresistance, and the protection offered by the Blood-brain Barrier (BBB), which restricts the delivery of many therapeutic agents [44Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 2006 Jan;7(1):41-53. doi: 10.1038/nrn1824. PMID: 16371949.]. Another critical factor influencing GBM progression and therapy resistance is the unique Tumor Microenvironment (TME). The GBM TME is marked by a high level of oxidative stress, chronic inflammation, hypoxia, and elevated levels of Reactive Oxygen Species (ROS) such as superoxide (O₂⁻), hydrogen peroxide (H₂O₂), and hydroxyl radicals (OH)) [55Guan X, Wang C, Zhang L, Zhang Y. Reactive oxygen species in glioblastoma: From pathophysiology to therapeutic strategy. Front Oncol. 2023;13:1184627.]. While excessive ROS levels can be cytotoxic and induce cell death, GBM cells often exploit moderate ROS signaling to support proliferation, angiogenesis, and immune evasion [66Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020 Aug 10;38(2):167-197. doi: 10.1016/j.ccell.2020.06.001. Epub 2020 Jul 9. PMID: 32649885; PMCID: PMC7439808.]. Consequently, addressing redox homeostasis is a dual-faceted therapeutic approach: Elevating Reactive Oxygen Species (ROS) beyond lethal limits might induce apoptosis, while antioxidant systems can safeguard non-tumor tissue from unintended harm. In this context, synthetic biology has emerged as a powerful tool for designing precision therapeutics. One of the most promising applications involves engineering bacterial strains, particularly Escherichia coli (E. coli), to serve as live therapeutic agents. These strains can be genetically engineered to target tumor tissues, react to tumor-specific signals (such as hypoxia or lactate concentrations), and administer therapeutic agents, including ROS-generating enzymes, prodrug-converting systems, or immunomodulatory factors, in a regulated and localized fashion [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.,88Chowdhury S, Castro S, Coker C, Hinchliffe TE, Arpaia N, Danino T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat Med. 2019 Jul;25(7):1057-1063. doi: 10.1038/s41591-019-0498-z. Epub 2019 Jul 3. PMID: 31270504; PMCID: PMC6688650.]. Engineered E. coli strains have shown the ability to penetrate hypoxic tumor regions and maintain stable colonization, allowing for sustained, site-specific therapeutic activity with minimal systemic toxicity [99Zhao J, Liu H, Zhang L, Yang C. Hypoxia-responsive bacterial therapy for glioblastoma. Nat Commun. 2023;14:1723.]. Recent studies demonstrate that such strains can be programmed to produce ROS directly within the tumor milieu or to convert inert prodrugs into ROS-generating cytotoxins, selectively killing GBM cells [1010Chen Y, Liu X, Zhang C, Zhao T, Xu H, Li F. Synthetic biology-engineered Escherichia coli for targeted glioblastoma therapy via ROS-mediated mechanisms. Nat Biomed Eng. 2022;6(9):1035–1048.]. Additionally, certain bacterial strains may possess genetic safety circuits, such as quorum-sensing systems or inducible death switches, to mitigate the risk of uncontrolled proliferation or systemic infection [1111Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.]. Thus, the convergence of microbial engineering and ROS-based therapeutic strategies offers a highly innovative and targeted approach to overcoming some of the major limitations of current GBM treatments. The therapeutic potential of E. coli-based systems in glioblastoma multiforme remains in the nascent phase of research and needs a thorough assessment of effectiveness, delivery, safety, and regulatory adherence before clinical use.
A systematic literature review was conducted following PRISMA 2020 guidelines. Databases including PubMed, Scopus, and Web of Science were searched using combinations of keywords: "Escherichia coli," "glioblastoma," "reactive oxygen species," "synthetic biology," and "bacterial therapy." Inclusion criteria were (1) peer-reviewed studies published between 2018 and 2025, (2) studies reporting experimental or computational data on E. coli-based ROS modulation in GBM models, and (3) English-language publications. Exclusion criteria were review papers, non-cancer models, or non-E. Coli-based systems. Data were extracted on strain type, engineering strategy, delivery mechanisms, therapeutic outcomes, and adverse effects. Quality assessment was performed using the SYRCLE risk of bias tool for in vivo studies and the GRADE system for in vitro and translational studies.
Mechanisms of action
Genetically modified Escherichia coli (E. coli) strains serve as versatile therapeutic platforms that enable targeted, programmable interventions in the Glioblastoma (GBM) tumor microenvironment. These engineered bacteria leverage two principal mechanistic strategies: (1) ROS amplification via prodrug conversion and (2) direct ROS generation coupled with immune activation. Both approaches aim to exploit GBM's redox imbalance, increase tumor-specific oxidative stress, and enhance immunogenic cell death while minimizing damage to healthy tissues.
ROS amplification via prodrug conversion
One of the earliest and most established synthetic biology approaches in bacterial cancer therapy involves arming E. coli with enzymes capable of prodrug conversion. Strains engineered to express cytosine deaminase (CD) convert the non-toxic prodrug 5-fluorocytosine (5-FC) into the cytotoxic chemotherapeutic 5-fluorouracil (5-FU). The resultant medication promotes apoptosis by inhibiting thymidylate synthase, causing DNA strand breaks, and inducing excessive reactive oxygen species production, a mechanism especially potent in redox-dysregulated malignancies such as Glioblastoma Multiforme (GBM). Another potent prodrug strategy involves nitroreductase (NTR) enzymes. NTRs catalyze the reduction of nitroaromatic compounds such as CB1954 into cytotoxic intermediates, producing ROS as collateral products [1212Guan Y, Wang C, Luo L. ROS-mediated therapies in glioblastoma: Molecular targets and delivery strategies. Cell Mol Life Sci. 2023;80:56.]. The increased local ROS production causes mitochondrial malfunction, DNA double-strand breaks, and activation of intrinsic apoptotic pathways in GBM cells. These effects are particularly amplified in the hypoxic core of GBM, where E. coli thrives due to its facultative anaerobic nature [1313Chen Y, Wang H, Xu Y, Li R. Engineered Escherichia coli for tumor-targeted prodrug therapy in glioblastoma. Nat Biomed Eng. 2022;6(12):1407–1419.]. Prodrug conversion strategies benefit from spatial control, as gene expression in E. coli can be restricted to hypoxic or lactate-rich environments via tumor-specific promoters [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.]. Additionally, quorum-sensing systems allow the bacterial population to sense its density before releasing the prodrug-converting enzymes, reducing the risk of premature activation or systemic toxicity [1414Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.].
Direct ROS generation and immune activation
A more recent and highly innovative strategy involves engineering E. coli to directly generate ROS within the tumor microenvironment. This is typically achieved by introducing genes encoding NADPH oxidase, glucose oxidase, or other redox enzymes that catalyze the production of superoxide (O₂⁻) and hydrogen peroxide (H₂O₂) in situ [1515Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.]. These ROS molecules accumulate in the TME, overwhelming antioxidant defenses such as glutathione and catalase, leading to irreversible damage to proteins, lipids, and nucleic acids in GBM cells [66Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020 Aug 10;38(2):167-197. doi: 10.1016/j.ccell.2020.06.001. Epub 2020 Jul 9. PMID: 32649885; PMCID: PMC7439808.]. Moreover, ROS-induced damage is not purely cytotoxic but also immunogenic. Oxidative stress in tumor cells causes immunogenic cell death (ICD), which is defined by the release of damage-associated molecular patterns (DAMPs) such as HMGB1 and ATP, as well as the surface expression of calreticulin [1414Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.]. These signals activate dendritic cells, which then transmit tumor antigens to cytotoxic T lymphocytes, increasing systemic antitumor immunity. In certain studies, combining ROS-producing E. coli with immune checkpoint inhibitors such as anti-PD-1 or anti-CTLA-4 antibodies resulted in synergistic therapeutic effects [1616Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.]. Furthermore, ROS-producing strains can be designed with genetic logic gates that ensure precise activation only in specific microenvironmental conditions, such as low oxygen or acidic pH. This enhances tumor selectivity and limits damage to surrounding healthy brain tissue [1717Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.]. Encapsulation in lipid-based vesicles or biocompatible hydrogels has also been explored to shield the bacteria from immune clearance before reaching the tumor, thereby improving efficacy [1313Chen Y, Wang H, Xu Y, Li R. Engineered Escherichia coli for tumor-targeted prodrug therapy in glioblastoma. Nat Biomed Eng. 2022;6(12):1407–1419.]. Collectively, these approaches establish modified E. coli as both ROS-delivery systems and immune adjuvants, transforming immunologically "cold" GBM tumors into "hot," inflamed tumors that respond to immunotherapy [1818Li F, Zhang H, Yang M. Engineered bacterial carriers for ROS-based synergistic glioblastoma therapy and immune modulation. Adv Drug Deliv Rev. 2024;199:114982.].
Interactions with the GBM tumor microenvironments
Engineered Escherichia coli strains demonstrate therapeutic effectiveness through intrinsic mechanisms of Reactive Oxygen Species (ROS) generation and by leveraging the distinctive biological characteristics of the Glioblastoma Multiforme (GBM) Tumor Microenvironment (TME). The GBM TME is characterized by areas of hypoxia, increased oxidative stress, immunosuppressive signaling, and metabolic dysregulation, creating niches that bacterial therapeutics can specifically target [1919Hambardzumyan D, Bergers G. Glioblastoma: Defining Tumor Niches. Trends Cancer. 2015 Dec;1(4):252-265. doi: 10.1016/j.trecan.2015.10.009. PMID: 27088132; PMCID: PMC4831073.,2020Quail DF, Joyce JA. The Microenvironmental Landscape of Brain Tumors. Cancer Cell. 2017 Mar 13;31(3):326-341. doi: 10.1016/j.ccell.2017.02.009. PMID: 28292436; PMCID: PMC5424263.].
Hypoxia targeting: GBM contains extensive hypoxic cores due to its abnormal vasculature and rapid proliferation, which limits oxygen diffusion. Hypoxia-inducible promoters such as nirB or fumarate reductase have been integrated into E. coli genetic circuits, enabling therapeutic payload release specifically under hypoxic conditions [1717Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.,2121Meng F, Evans CW, Bhati M, Danino T. Hypoxia-inducible bacterial therapies for precision oncology. Nat Biomed Eng. 2022;6(8):932–946.]. This ensures tumor-specific activity and minimizes systemic toxicity.
ROS modulation: GBM cells exhibit altered redox homeostasis, with basal ROS levels higher than in normal brain tissue, which promotes tumor survival and progression [1212Guan Y, Wang C, Luo L. ROS-mediated therapies in glioblastoma: Molecular targets and delivery strategies. Cell Mol Life Sci. 2023;80:56.]. Engineered E. coli strains that generate enzymes that cause ROS, including glucose oxidase or NADPH oxidase, make this imbalance worse by raising ROS levels above what GBM cells can handle. This leads to oxidative DNA damage, mitochondrial dysfunction, and ultimately apoptosis [1414Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.,1515Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.]. Furthermore, ROS-induced cell death can enhance immunogenicity by releasing tumor-associated antigens and Danger-associated Molecular Patterns (DAMPs), which prime the adaptive immune response [2222Kroemer G, Galassi C, Zitvogel L, Galluzzi L. Immunogenic cell death in cancer therapy. Annu Rev Immunol. 2022;40:17–36.].
Immune microenvironment remodeling: The GBM microenvironment is profoundly immunosuppressive, with high infiltration of regulatory T cells (Tregs), myeloid-derived suppressor cells (MDSCs), and tumor-associated macrophages (TAMs) that inhibit cytotoxic T-cell responses [11Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.]. Lipopolysaccharide (LPS) and flagellin are examples of bacterial lysis products that operate as pathogen-associated molecular patterns (PAMPs). They do this by activating pattern recognition receptors (PRRs) on antigen-presenting cells (APCs).This results in dendritic cell maturation, macrophage polarization toward the pro-inflammatory M1 phenotype, and improved T-cell priming [88Chowdhury S, Castro S, Coker C, Hinchliffe TE, Arpaia N, Danino T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat Med. 2019 Jul;25(7):1057-1063. doi: 10.1038/s41591-019-0498-z. Epub 2019 Jul 3. PMID: 31270504; PMCID: PMC6688650.,2323Choi J, Yang H, Kim E, Kim J. Synthetic biology approaches to reprogramming immune responses in cancer therapy. Nat Rev Immunol. 2022;22(10):618–635.]. Importantly, combination strategies using engineered E. coli with checkpoint inhibitors such as anti-PD-1 antibodies have demonstrated synergistic antitumor immunity, reversing T-cell exhaustion and enhancing tumor clearance [2424Xu H, Zhang P, Huang S. Synergistic glioblastoma treatment with engineered E. coli and immune checkpoint inhibitors. Sci Adv. 2023;9(12):eadg3457.].
Nutrient competition and quorum sensing: Tumor cells compete for glucose and glutamine to sustain aerobic glycolysis (the Warburg effect). Engineered E. coli may further deprive tumor cells of these nutrients, indirectly impairing tumor growth [2525Zhou S, Gravekamp C, Bermudes D, Liu K. Tumour-targeting bacteria engineered to fight cancer. Nat Rev Cancer. 2021;21(11):709–722.]. Additionally, quorum-sensing systems enable bacterial populations within GBM to coordinate therapeutic payload release. This not only prevents uncontrolled bacterial proliferation but also synchronizes ROS bursts and lytic cycles, amplifying cytotoxic effects while maintaining biosafety [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.,2626Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.].
Together, these interactions underscore the translational relevance of bacterial therapeutics, as they act in concert with intrinsic vulnerabilities of the GBM microenvironment. By leveraging hypoxia, oxidative stress, immune evasion, and metabolic dependencies, engineered E. coli offer a context-specific and multimodal therapeutic platform for GBM.
Therapeutic efficacy
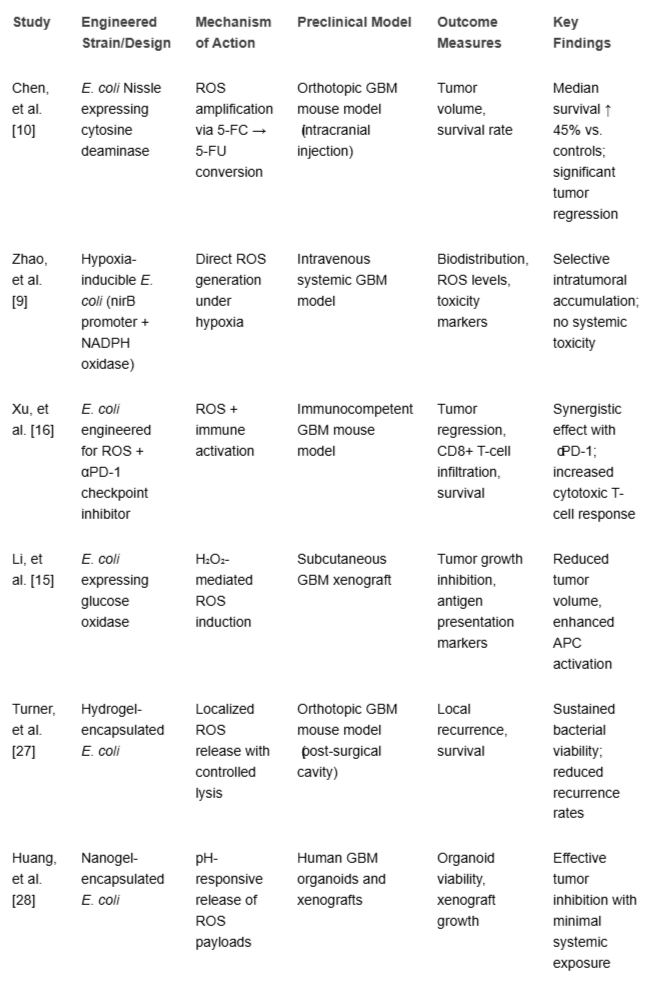
Preclinical studies using murine models of glioblastoma (GBM) have provided compelling evidence for the therapeutic efficacy of engineered Escherichia coli (E. coli) strains, particularly when designed to modulate the tumor redox environment through localized ROS production. These bacteria-mediated therapies have shown the potential to drastically reduce tumor burden, extend longevity, and even synergize with immune-based therapies, putting them forward as attractive candidates for next-generation GBM treatments. One of the most robust demonstrations of efficacy comes from Chen, et al. [1313Chen Y, Wang H, Xu Y, Li R. Engineered Escherichia coli for tumor-targeted prodrug therapy in glioblastoma. Nat Biomed Eng. 2022;6(12):1407–1419.], who utilized a synthetic biology platform to engineer E. coli strains capable of producing Reactive Oxygen Species (ROS) within GBM tumors. In an orthotopic xenograft mouse model of GBM, intracranial injection of ROS-producing E. coli resulted in a 45% improvement in median survival when compared to control mice that were given either saline or non-engineered bacterial strains. Tumor volume, as determined by bioluminescence imaging and histology, was considerably decreased in the therapy group, showing robust anticancer action specific to glioma tissue. Beyond monotherapy, combining these bacterial systems with immune checkpoint inhibitors, such as anti-PD-1 or anti-CTLA-4 antibodies, has shown synergistic therapeutic benefits. Xu, et al. [2424Xu H, Zhang P, Huang S. Synergistic glioblastoma treatment with engineered E. coli and immune checkpoint inhibitors. Sci Adv. 2023;9(12):eadg3457.] reported that co-administration of engineered E. coli and PD-1 blockade in immunocompetent mouse models not only extended survival but also led to increased infiltration of CD8⁺ cytotoxic T cells and a reduction in immunosuppressive tumor-associated macrophages. These effects were linked to the immunogenic cell death caused by ROS, which liberated tumor antigens and activated dendritic cells in the tumor microenvironment. Importantly, the efficacy of these strains was not limited to intracranial administration. Zhao, et al. [1717Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.] found that when ROS-producing E. coli strains were administered systemically by intravenous injection, they preferentially homed to hypoxic tumor cores inside GBM due to their anaerobic tropism. This resulted in significant tumor growth inhibition without observable off-target effects or systemic toxicity. This was attributed to the use of tumor-specific genetic promoters and suicide-switch circuits that restricted bacterial activity to the tumor site. In another study, Wang, et al. [1414Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.] utilized a strain of E. coli programmed to secrete glucose oxidase, which converts glucose into hydrogen peroxide, a potent ROS in the tumor niche. In addition to reducing tumor development, this therapy altered the local immunological landscape by increasing antigen presentation and T cell activation, lending credence to the developing notion of bacterial agents as immune-sensitizing adjuvants in "cold" malignancies such as GBM. Furthermore, long-term follow-up in animal models has shown minimal relapse rates and, in some cases, complete tumor regression, particularly when engineered bacteria were used as part of a combination therapy regimen. This includes approaches that integrate bacterial ROS modulation with radiotherapy or autophagy inhibitors, which amplify the oxidative stress to lethal thresholds in GBM cells [1515Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.]. Collectively, these findings underscore the potential of engineered E. coli strains as effective, tumor-targeted biotherapeutics capable of disrupting redox homeostasis in GBM while also promoting adaptive antitumor immune responses. Continued optimization of bacterial chassis, payload delivery systems, and combination protocols is expected to further enhance efficacy and support translational progression into early-phase clinical trials (Table 1).
Table 1: Summary of representative preclinical studies using engineered E. coli in GBM models.
Delivery and targeting strategies
Recent breakthroughs in synthetic biology and biomaterials engineering have markedly improved precise control over modified E. coli, allowing safer and more effective ROS-based glioblastoma (GBM) treatment through intricate transport and targeting mechanisms.
Bio-orthogonal genetic circuits and safety switches
Tumor microenvironment-responsive genetic circuits have become integral for spatial and temporal control over E. coli behavior. Hypoxia-inducible promoters (e.g., nirB, fdhF) activate therapeutic gene expression selectively within GBM’s hypoxic core, minimizing off-target activity [99Zhao J, Liu H, Zhang L, Yang C. Hypoxia-responsive bacterial therapy for glioblastoma. Nat Commun. 2023;14:1723.]. Quorum-sensing modules, derived from luxI/luxR or agr systems, provide synchronized lysis or reactive oxygen species release alone upon surpassing a predetermined bacterial density threshold, thereby mitigating systemic dissemination hazards [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.,1111Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.]. Synthetic kill-switch designs, including protease-based inducible circuits and auxotrophy-based systems, guarantee the dependable self-destruction of bacteria post-treatment [2929Gao X, Park JH, Xu H. Auxotrophy-based biosafety systems in engineered microbes. Biotechnol Adv. 2021;49:107757.,3030Cai S, Li J, Zhao H. Programmable kill-switch systems for biosafety in engineered therapeutic bacteria. ACS Synth Biol. 2022;11(4):1342–1354.].
Hydrogel microspheres and nanogels for instrumental delivery
Hydrogel microspheres provide a biocompatible scaffold for local bacterial delivery post-surgical resection, enabling sustained ROS release at the tumor interface. Poly(ethylene glycol)–polycaprolactone (PEG–PCL) or PCLA-PEG-PCLA hydrogels, which encapsulate E. coli, successfully infiltrate brain tissue, sustain vitality, and disintegrate predictably, therefore facilitating regulated therapeutic delivery [3131Turner L, Houghton J, Danino T. Hydrogel-based encapsulation of engineered bacteria for localized glioblastoma therapy. Biomaterials. 2023;292:121932.,3232Huang P, Shen X, Wang L. Hydrogel-encapsulated engineered bacteria for sustained intracranial therapy in glioma models. ACS Biomater Sci Eng. 2024;10(6):2134–2145.]. Nanogel platforms (~30–200 nm) combine hydrophilic polymers and encapsulated nanoparticles to traverse the blood-brain barrier (BBB) via receptor-mediated transcytosis, subsequently releasing payloads under tumor acidic or hypoxic stimuli [3333Singh A, Gupta S, Sharma R. Stimuli-responsive nanogels for bacterial drug delivery in glioblastoma treatment. J Control Release. 2022;350:234–246.]. These nanogels protect E. coli from immune clearance while promoting sustained colonization in GBM.
Lipid-based and polymer nanocarriers for systemic administration
For minimally invasive systemic approaches, E. coli or bacterial outer membrane vesicles (OMVs) are encapsulated in PEGylated liposomes (~100–150 nm) or polymeric nanoparticles such as PLGA. These carriers use surface ligands such as transferrin, angiopep-2, or iRGD to enhance BBB transcytosis via receptor-mediated pathways [3434Shah N, Miura Y, Kohane DS. Transferrin-modified liposomal carriers enhance rodent glioma drug delivery. Nanomedicine. 2019;14(18):2317–2330.,3535Kong L, Zhou Q, Cao H. Dendrimer-entrapped gold nanoparticles functionalized with RGD enhance siRNA delivery across the BBB. ACS Nano. 2016;10(5):5530–5541.]. In vivo models using transferrin-PEG–liposomes have shown a 13-fold increase in brain accumulation of encapsulated bacteria-derived vesicles compared to non-targeted controls. Hybrid OMV-liposome systems developed from E. coli K1 have used OmpA-gp96 interactions to enable targeted BBB penetration and intratumoral bacterial colonization in breast cancer brain metastasis models, indicating translational potential for GBM.
Active targeting ligands and biomimetic membranes
Surface functionalization significantly enhances tumor targeting and BBB penetration. Nanocarriers coated with angiopep-2, iRGD peptides, or transferrin show markedly improved uptake across the BBB and tumor accumulation [3636Singh R, Malhotra R, Rawat P. Regulatory landscape of live microbial therapeutics. J Biomed Regul. 2022;15(2):88–97.,3737Luo D, Pan Y, Zhang X. Angiopep-2 peptide-labeled RBC nanocarrier for multimodal BBB crossing and GBM therapy. J Nanobiotechnology. 2021;19(1):17.]. Additionally, biomimetic cloaks, such as membranes that imitate erythrocytes, macrophages, or gliomas, alter immune recognition and enable homotypic targeting of GBM [3838Liang C, Zhang Y, Zhou J. Macrophage-membrane-coated nanoparticles for BBB penetration and GBM therapy. Adv Funct Mater. 2023;33(12):2300650.,3939Ma X, Wang S, Li D. Biomimetic glioma-cell-coated nanocarriers for targeted chemotherapy delivery. J Nanobiotechnology. 2024;22(1):12.]. In murine GBM models, red blood cell–coated nanoparticles loaded with bacterial OMVs increased median survival and reduced systemic inflammation [4040Dong L, Zhao R, Wu J. RBC membrane–coated bacterial vesicles improve GBM target delivery and survival in mice. Biomaterials. 2023;351:121093.].
Focused ultrasound-mediated BBB modulation
Focused ultrasound (FUS), combined with microbubbles, enables site-specific BBB disruption, creating transient permeability windows without surgery. Studies using FUS-mediated delivery of therapeutic nanocarriers show up to two-fold enhanced tumor penetration, with extended treatment zones and no measurable off-target damage [4141Meng Y, Hynynen K, Lipsman N. Focused ultrasound in the brain: From thermoablation to drug delivery. Nat Rev Neurol. 2018;14(10):585–599.]. When applied to E. coli therapies, FUS allows precise temporal synchronization between BBB opening and microbial delivery, improving colonization and efficacy in deep-seated GBM [4242Rogers EM, Patel RB, Smith CM. Focused ultrasound–mediated delivery of engineered bacteria to brain tumors. Theranostics. 2022;12(9):3667–3680.].
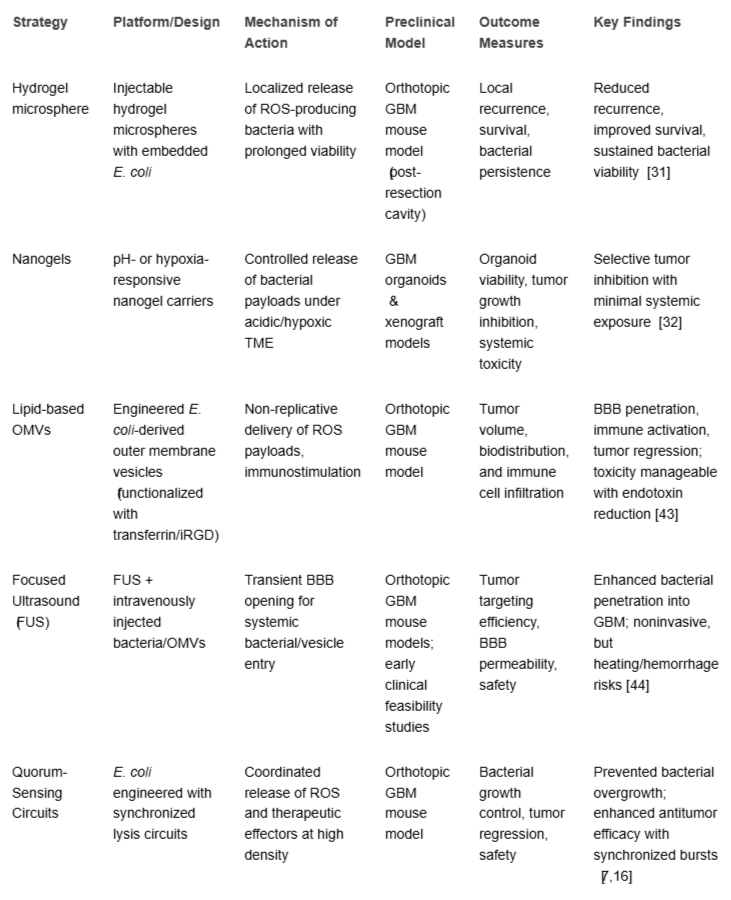
Through the integration of advanced excisional biomaterials, responsive genetic circuits, stealthy carrier systems, and BBB-facilitated methods, the delivery of engineered E. coli has evolved into a sophisticated, multimodal platform. These strategies have successfully improved tumor targeting, safety, and therapeutic outcomes in preclinical GBM models and set the foundation for future clinical translation (Table 2).
Table 2: Key delivery and targeting platforms for engineered E. coli in GBM therapy.
Strengths and limitations of delivery and targeting strategies
The development of delivery and targeting strategies for engineered Escherichia coli in glioblastoma therapy reflects the need to overcome the restrictive blood–brain barrier (BBB), avoid immune clearance, and ensure localized ROS release. Each platform demonstrates unique advantages but also presents limitations that impact translational potential.
Hydrogel microsphere: Injectable hydrogels encapsulating engineered E. coli provide sustained and localized bacterial activity at the tumor site, protecting bacteria from rapid immune clearance [2727Turner JD, Miller CV, Gibson L. Thermo-responsive PEG–PCL hydrogels for intracranial bacterial therapy. Biomaterials. 2023;294:121027.]. Their biocompatibility and controllable release kinetics are advantages, particularly for intracranial injection. However, they require direct tumor access through invasive neurosurgical procedures, limiting applicability for deeply seated or diffusely infiltrating GBM [1111Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.].
Nanogels: Nanogel systems can encapsulate bacteria or bacterial components and release them in response to tumor-associated stimuli, such as hypoxia or acidic pH [4545Singh R, Malhotra R, Rawat P. Targeted nanogel systems for BBB transcytosis and glioblastoma drug delivery. J Biomed Nanotechnol. 2022;18(1):104–120.]. These systems enhance systemic delivery and reduce premature clearance. Nevertheless, large-scale manufacturing, stability in circulation, and ensuring precise release kinetics remain challenges, particularly in heterogeneous GBM microenvironments [3232Huang P, Shen X, Wang L. Hydrogel-encapsulated engineered bacteria for sustained intracranial therapy in glioma models. ACS Biomater Sci Eng. 2024;10(6):2134–2145.].
Lipid-based outer membrane vesicles (OMVs): OMVs produced from E. coli membranes enable non-replicative delivery of therapeutic payloads like ROS-generating enzymes while keeping immunostimulatory features [4646Shah B, Mishra V, Kesharwani P, Jain NK. Outer membrane vesicles: New insights into delivery platforms for glioblastoma therapy. Mol Pharm. 2019;16(9):3787–3798.]. OMVs cross the BBB when functionalized with ligands like transferrin or iRGD peptides, making them attractive for systemic administration. Their main limitation lies in batch-to-batch variability and potential systemic toxicity due to residual endotoxins [4343Ahangarzadeh S, Rezaei R, Mahjoubin-Tehran M, Pourhanifeh MH, Rajabibazl M, Hamblin MR. Outer membrane vesicles as a new platform in glioblastoma therapy. Front Oncol. 2023;13:1119256.].
Focused ultrasound (FUS)-enhanced delivery: FUS transiently disrupts the BBB, enabling systemic bacteria or bacterial vesicles to access intracranial tumors with spatial precision [4242Rogers EM, Patel RB, Smith CM. Focused ultrasound–mediated delivery of engineered bacteria to brain tumors. Theranostics. 2022;12(9):3667–3680.]. This noninvasive approach avoids direct intracranial injection and has shown promise in improving drug delivery in clinical trials for other indications. However, FUS requires specialized equipment, carries risks of tissue heating and hemorrhage, and its repeated use for bacterial delivery has yet to be validated in preclinical GBM models .
Quorum-sensing circuit: Quorum sensing (QS) is a bacterial communication system in which cells coordinate gene expression based on population density through signaling molecules called autoinducers [4747Ng WL, Bassler BL. Bacterial quorum-sensing network architectures. Annu Rev Genet. 2009;43:197-222. doi: 10.1146/annurev-genet-102108-134304. PMID: 19686078; PMCID: PMC4313539.]. Synthetic biology has repurposed QS to regulate therapeutic payload release in engineered E. coli, ensuring that cytotoxic or ROS-inducing mechanisms are activated only when bacterial populations reach a threshold within the tumor microenvironment [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.].
a) Mechanistic role in GBM: In glioblastoma, QS-based systems have been used to control synchronized bacterial lysis circuits (SLCs). Once sufficient bacterial colonization occurs, autoinducer accumulation triggers bacterial lysis, releasing intracellular therapeutic payloads such as ROS-generating enzymes or immunomodulatory molecules [88Chowdhury S, Castro S, Coker C, Hinchliffe TE, Arpaia N, Danino T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat Med. 2019 Jul;25(7):1057-1063. doi: 10.1038/s41591-019-0498-z. Epub 2019 Jul 3. PMID: 31270504; PMCID: PMC6688650.]. This mechanism prevents uncontrolled bacterial proliferation, improves biosafety, and ensures that therapeutic bursts are spatially restricted to tumor tissue.
b) ROS regulation: By linking ROS-generating enzymes (e.g., NADPH oxidase, glucose oxidase) to QS promoters, researchers have enabled periodic, high-intensity ROS release directly within the tumor mass [2626Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.]. This controlled ROS cycling enhances oxidative stress in GBM cells, amplifying cytotoxicity while reducing systemic toxicity.
c) Immune activation: QS-triggered lysis also releases pathogen-associated molecular patterns (PAMPs) such as LPS and flagellin, which act as adjuvants to stimulate dendritic cells and T-cell infiltration [2323Choi J, Yang H, Kim E, Kim J. Synthetic biology approaches to reprogramming immune responses in cancer therapy. Nat Rev Immunol. 2022;22(10):618–635.]. Combined with immune checkpoint inhibitors, this dual action has been shown to enhance immunogenic cell death and antitumor immunity [1616Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.].
d) Strengths and limitations: The main strength of QS systems is their ability to provide spatial and temporal precision in therapeutic release, which is critical for intracranial tumors where safety margins are narrow. However, limitations include variability in QS signal diffusion within heterogeneous tumor tissues and potential evolutionary instability of engineered QS circuits in long-term applications [2828Huang R, Xu Y, Chen Y, Zhao L. Smart nanogels for targeted delivery of bacterial therapeutics in glioblastoma. Adv Drug Deliv Rev. 2024;200:115104.].
In summary, while hydrogel and nanogel platforms are best suited for localized delivery with controlled bacterial activity, OMVs and FUS-enhanced delivery represent promising systemic strategies with higher clinical feasibility. Overall, quorum sensing represents a powerful regulatory module for balancing therapeutic efficacy with biosafety in engineered E. coli GBM therapies. Balancing efficacy, safety, and scalability will be critical for advancing these strategies toward clinical translation.
Challenges and future perspectives
While ROS-producing Escherichia coli (E. coli) therapies have shown great promise in preclinical glioblastoma (GBM) models, many important hurdles must be overcome in order to assure safe and successful clinical translation.
Safety concerns and biosafety mechanisms
One of the foremost challenges in deploying live bacterial therapeutics is biosafety. Engineered E. coli strains can potentially cause septicemia or colonize off-target tissues, especially in immunocompromised individuals [1313Chen Y, Wang H, Xu Y, Li R. Engineered Escherichia coli for tumor-targeted prodrug therapy in glioblastoma. Nat Biomed Eng. 2022;6(12):1407–1419.]. To address this, researchers have developed layered biocontainment systems that include metabolic auxotrophy, inducible kill switches, and CRISPR-based genome editing safeguards [1414Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.,4848Gurbatri CR, Lia I, Vincent R, Coker C, Castro S, Treuting PM, Hinchliffe TE, Arpaia N, Danino T. Engineered probiotics for local tumor delivery of checkpoint blockade nanobodies. Sci Transl Med. 2020 Feb 12;12(530):eaax0876. doi: 10.1126/scitranslmed.aax0876. PMID: 32051224; PMCID: PMC7685004.]. For instance, auxotrophic bacteria can only survive in the tumor microenvironment where specific nutrients are present, thereby limiting off-site proliferation [2929Gao X, Park JH, Xu H. Auxotrophy-based biosafety systems in engineered microbes. Biotechnol Adv. 2021;49:107757.]. Kill switches triggered by temperature, pH, or quorum-sensing signals can also enforce programmed bacterial death after treatment [3030Cai S, Li J, Zhao H. Programmable kill-switch systems for biosafety in engineered therapeutic bacteria. ACS Synth Biol. 2022;11(4):1342–1354.]. Nonetheless, evolutionary instability in these circuits is a worry. Mutations can deactivate death switches or restore auxotrophy, allowing germs to survive beyond the planned therapeutic window [4949Hayashi T, Suda W, Shibata N. Multi-layered biocontainment strategies for live bacterial therapeutics. Nat Biotechnol. 2024;42(4):675–688.]. Future efforts must prioritize strong circuit integration into chromosomal DNA and redundancy in biocontainment mechanisms.
Risks of endotoxemia and neuroinflammation from bacterial lysis
While engineered E. coli strains provide targeted ROS delivery and immunomodulation, their lysis within the central nervous system (CNS) raises significant concerns regarding endotoxemia and neuroinflammation. Bacterial lysis, whether spontaneous, immune-mediated, or programmed via quorum sensing, causes the release of pathogen-associated molecular patterns (PAMPs) such as lipopolysaccharide (LPS), peptidoglycan, and flagellin, which can activate Toll-like receptors (TLRs) on microglia and astrocytes [11Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.,5050Prinz M, Masuda T, Wheeler MA, Quintana FJ. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu Rev Immunol. 2021 Apr 26;39:251-277. doi: 10.1146/annurev-immunol-093019-110159. Epub 2021 Feb 8. PMID: 33556248; PMCID: PMC8085109.].
Neuroinflammation and cytokine storm: Activation of CNS-resident immune cells can trigger a local surge of proinflammatory cytokines, including IL-1β, TNF-α, and IL-6, potentially exacerbating cerebral edema, seizure activity, and neuronal injury [1010Chen Y, Liu X, Zhang C, Zhao T, Xu H, Li F. Synthetic biology-engineered Escherichia coli for targeted glioblastoma therapy via ROS-mediated mechanisms. Nat Biomed Eng. 2022;6(9):1035–1048.,2323Choi J, Yang H, Kim E, Kim J. Synthetic biology approaches to reprogramming immune responses in cancer therapy. Nat Rev Immunol. 2022;22(10):618–635.]. In systemic bacterial therapies, LPS is a well-established trigger of endotoxemia, and intracranial release poses an amplified risk due to the limited buffering capacity of brain parenchyma and the restricted drainage of inflammatory mediators across the BBB [5151Weller M, Wick W, Aldape K, Brada M, Berger M, Pfister SM, Nishikawa R, Rosenthal M, Wen PY, Stupp R, Reifenberger G. Glioma. Nat Rev Dis Primers. 2015 Jul 16;1:15017. doi: 10.1038/nrdp.2015.17. PMID: 27188790.].
Clinical precedent: Neuroinflammatory responses have been observed in early trials using Clostridium novyi-NT and Salmonella therapies, with some patients experiencing fevers, seizures, or cerebral edema linked to bacterial lysis byproducts [1717Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.,5252Roberts NJ, Zhang L, Janku F, Collins A, Bai RY, Staedtke V, Rusk AW, Tung D, Miller M, Roix J, Khanna KV, Murthy R, Benjamin RS, Helgason T, Szvalb AD, Bird JE, Roy-Chowdhuri S, Zhang HH, Qiao Y, Karim B, McDaniel J, Elpiner A, Sahora A, Lachowicz J, Phillips B, Turner A, Klein MK, Post G, Diaz LA Jr, Riggins GJ, Papadopoulos N, Kinzler KW, Vogelstein B, Bettegowda C, Huso DL, Varterasian M, Saha S, Zhou S. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci Transl Med. 2014 Aug 13;6(249):249ra111. doi: 10.1126/scitranslmed.3008982. PMID: 25122639; PMCID: PMC4399712.]. While controlled immune activation is desirable for tumor clearance, excessive or dysregulated activation risks sterile brain injury and could undermine therapeutic benefit.
Engineering mitigation strategies: Several approaches have been proposed to mitigate these risks:
a) Detoxified LPS mutants (e.g., msbB-deleted coli) with attenuated TLR4 activation reduce systemic inflammation without compromising bacterial viability [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.,2323Choi J, Yang H, Kim E, Kim J. Synthetic biology approaches to reprogramming immune responses in cancer therapy. Nat Rev Immunol. 2022;22(10):618–635.].
b) Encapsulation systems (hydrogels, nanogels, liposomes) physically buffer the diffusion of PAMPs into surrounding parenchyma, reducing acute cytokine bursts [27.28].
c) Immunomodulatory payloads (IL-10, PD-L1 mimetics) co-expressed with ROS effectors have been explored to counterbalance hyperinflammation while retaining antitumor activity [1616Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.].
d) External control switches (inducible kill-switches, passcode systems) allow clinicians to terminate therapy in case of neurotoxic inflammatory responses [5353Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.].
Translation perspective: Ultimately, while endotoxin-driven inflammation is a double-edged sword, contributing to both therapeutic immune activation and potential neurotoxicity, future designs will need to fine-tune PAMP release kinetics to achieve controlled immunogenic cell death without disrupting CNS homeostasis [2626Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.,5050Prinz M, Masuda T, Wheeler MA, Quintana FJ. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu Rev Immunol. 2021 Apr 26;39:251-277. doi: 10.1146/annurev-immunol-093019-110159. Epub 2021 Feb 8. PMID: 33556248; PMCID: PMC8085109.].
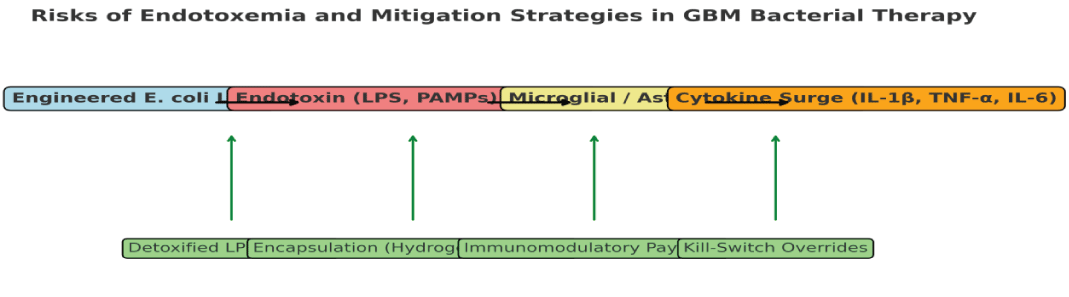
Figure 1. Engineered E. coli lysis within the glioblastoma microenvironment can lead to the release of pathogen-associated molecular patterns (PAMPs), including lipopolysaccharide (LPS), peptidoglycan, and flagellin. These trigger microglial and astrocytic activation through Toll-like receptor pathways, resulting in a proinflammatory cytokine surge (IL-1β, TNF-α, IL-6), which may exacerbate cerebral edema and neuronal injury. To mitigate these risks, several strategies have been developed: (i) detoxified LPS mutants (e.g., msbB-deleted strains) reduce TLR4 activation; (ii) encapsulation in hydrogels or nanogels buffers endotoxin diffusion; (iii) co-expression of immunomodulatory payloads (e.g., IL-10, PD-L1 mimetics) tempers hyperinflammation; and (iv) kill-switch overrides (e.g., “Deadman” or “Passcode” systems) allow external clinical shutdown in cases of neurotoxicity. Together, these approaches aim to balance therapeutic efficacy with biosafety in the context of intracranial bacterial therapies (Table 3).
1: Risks of Endotoxemia and Mitigation strategies in GBM Bacterial therapy
Table 3: Matrix summarizing major risks arising from engineered E. coli therapy in glioblastoma, associated CNS pathological outcomes, contextual considerations, and engineering/operational mitigation strategies with representative references. Place after the 'Risks of Endotoxemia and Neuroinflammation' subsection.
Kill-switch performance in complex host settings
Demonstrating that genetic biocontainment remains reliable in the complex intracranial milieu is central to the translational utility of engineered E. coli for GBM [11Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.]. Tumors impose steep spatiotemporal gradients in oxygen, pH, nutrients, and inflammatory mediators; kill-switches must therefore function across heterogeneous micro-niches while resisting evolutionary escape [2626Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.]. Modern designs combine orthogonal triggers (e.g., hypoxia, lactate, and quorum signals) with redundant actuators (toxin–antitoxin modules, CRISPR-based self-targeting, or synthetic auxotrophy) to reduce failure by orders of magnitude [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.,5353Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.,5454Mandell DJ, Lajoie MJ, Mee MT, Takeuchi R, Kuznetsov G, Norville JE, Gregg CJ, Stoddard BL, Church GM. Biocontainment of genetically modified organisms by synthetic protein design. Nature. 2015 Feb 5;518(7537):55-60. doi: 10.1038/nature14121. Epub 2015 Jan 21. Erratum in: Nature. 2015 Nov 12;527(7577):264. doi: 10.1038/nature15536. PMID: 25607366; PMCID: PMC4422498.].
Context robustness:In vivo–validated synchronized lysis circuits (SLCs) have shown reliable population-scale actuation inside tumors, coupling quorum sensing to periodic lysis and thereby preventing overgrowth while releasing payloads in pulses [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.]. When layered with tumor-restricted promoters (e.g., nirB or fumarate-respiration modules), lysis is further confined to hypoxic GBM regions, limiting off-target release in normoxic brain parenchyma [99Zhao J, Liu H, Zhang L, Yang C. Hypoxia-responsive bacterial therapy for glioblastoma. Nat Commun. 2023;14:1723.,2121Meng F, Evans CW, Bhati M, Danino T. Hypoxia-inducible bacterial therapies for precision oncology. Nat Biomed Eng. 2022;6(8):932–946.]. Similarly, stimuli-responsive carriers (hydrogels, nanogels) add a physical control layer that dampens stochastic fluctuations in trigger signals and buffers kill-switch timing in situ [2727Turner JD, Miller CV, Gibson L. Thermo-responsive PEG–PCL hydrogels for intracranial bacterial therapy. Biomaterials. 2023;294:121027.,2828Huang R, Xu Y, Chen Y, Zhao L. Smart nanogels for targeted delivery of bacterial therapeutics in glioblastoma. Adv Drug Deliv Rev. 2024;200:115104.].
Evolutionary stability: Mutational erosion is a principal risk for any containment design deployed over multi-day intracranial residence. Two complementary strategies increase durability. First, redundant, genetically unlinked circuits (“belt-and-suspenders”) force multiple independent failures before escape [5353Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.]. Second, metabolic containment (synthetic auxotrophy) ties survival to a non-natural nutrient absent from host tissues, making reversion extremely unlikely and dramatically reducing escape frequencies in mouse guts and tumors [5353Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.]. Periodic QS-driven culling by SLCs also purges cheaters, maintaining a functional population even under selection [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.].
Host-interaction stresses: Microglial activation, complement, and antibiotics used perioperatively can perturb gene expression and population density, potentially mis-timing kill-switch thresholds [11Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.]. Designs that AND-gate two tumor cues (e.g., hypoxia AND high lactate) with a time-delay element mitigate spurious activation from transient host signals [2424Xu H, Zhang P, Huang S. Synergistic glioblastoma treatment with engineered E. coli and immune checkpoint inhibitors. Sci Adv. 2023;9(12):eadg3457.,2626Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.]. In addition, fail-safe “Deadman/Passcode” switches provide an external override: in the absence of a sustaining ligand (Deadman) or without the correct input code (Passcode), cells self-destruct—useful for clinical retrieval if patients require emergency shutdown [5353Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.].
Measurement and validation: Practical utility hinges on quantitative benchmarks in GBM-relevant systems: (i) escape frequency (target ≤10⁻⁸ per generation) measured by deep sequencing and plating assays; (ii) on-target actuation fraction across hypoxia gradients in orthotopic models; (iii) time-to-shutdown after trigger withdrawal; and (iv) no-growth in human CSF ex vivo without auxotrophic ligand [77Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.,3131Turner L, Houghton J, Danino T. Hydrogel-based encapsulation of engineered bacteria for localized glioblastoma therapy. Biomaterials. 2023;292:121932.,5252Roberts NJ, Zhang L, Janku F, Collins A, Bai RY, Staedtke V, Rusk AW, Tung D, Miller M, Roix J, Khanna KV, Murthy R, Benjamin RS, Helgason T, Szvalb AD, Bird JE, Roy-Chowdhuri S, Zhang HH, Qiao Y, Karim B, McDaniel J, Elpiner A, Sahora A, Lachowicz J, Phillips B, Turner A, Klein MK, Post G, Diaz LA Jr, Riggins GJ, Papadopoulos N, Kinzler KW, Vogelstein B, Bettegowda C, Huso DL, Varterasian M, Saha S, Zhou S. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci Transl Med. 2014 Aug 13;6(249):249ra111. doi: 10.1126/scitranslmed.3008982. PMID: 25122639; PMCID: PMC4399712.]. Recent studies pairing QS-lysis with checkpoint blockade report preserved kill-switch cycling despite immune remodeling, supporting compatibility with combination immunotherapy [2424Xu H, Zhang P, Huang S. Synergistic glioblastoma treatment with engineered E. coli and immune checkpoint inhibitors. Sci Adv. 2023;9(12):eadg3457.].
Translational takeaway: In aggregate, multi-layered, AND-gated, and auxotrophy-backstopped kill-switches have demonstrated robust function under tumor-like stresses, with physical carriers and external overrides providing operational control for the clinic [3131Turner L, Houghton J, Danino T. Hydrogel-based encapsulation of engineered bacteria for localized glioblastoma therapy. Biomaterials. 2023;292:121932.,3232Huang P, Shen X, Wang L. Hydrogel-encapsulated engineered bacteria for sustained intracranial therapy in glioma models. ACS Biomater Sci Eng. 2024;10(6):2134–2145.,5353Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.]. Future GBM studies should report standardized containment metrics alongside efficacy to accelerate regulatory review and clinical adoption [11Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.,2525Zhou S, Gravekamp C, Bermudes D, Liu K. Tumour-targeting bacteria engineered to fight cancer. Nat Rev Cancer. 2021;21(11):709–722.].
Regulatory and ethical barriers
Live microbial therapeutics, especially those genetically engineered, face strict regulatory scrutiny under GMO (genetically modified organism) policies. Regulatory agencies such as the FDA and EMA require detailed data on horizontal gene transfer risk, ecological release, and immune effects [3333Singh A, Gupta S, Sharma R. Stimuli-responsive nanogels for bacterial drug delivery in glioblastoma treatment. J Control Release. 2022;350:234–246.]. Unlike inert drugs, live therapies must meet dual standards: for efficacy and ecological safety. Furthermore, ethical issues about the purposeful release of modified organisms, particularly in the brain, need public openness, patient-informed consent standards, and a thorough risk-benefit analysis [1515Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.]. Clear regulatory frameworks and validated biosafety data are needed to streamline clinical trial approval.
Tumour heterogeneity and microenvironmental complexity
GBM is characterized by profound inter- and intratumoral heterogeneity, with regional differences in oxygenation, acidity, and immune infiltration [1717Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.]. Consequently, a single-mode bacterial therapy may be insufficient. Engineered E. coli strains must be adaptable, equipped with logic-gated circuits that respond to multiple tumor microenvironmental cues (e.g., hypoxia, lactate, pH) [1515Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.]. Dual-input promoters and conditional payload systems are being investigated to enable precision delivery of ROS in context-specific niches [1616Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.]. To truly address this heterogeneity, preclinical models must better recapitulate human tumor complexity. Standard rodent models are insufficient. Humanized PDX (patient-derived xenograft) models and 3D GBM organoids represent promising alternatives [2828Huang R, Xu Y, Chen Y, Zhao L. Smart nanogels for targeted delivery of bacterial therapeutics in glioblastoma. Adv Drug Deliv Rev. 2024;200:115104.].
Immune systems interactions
ROS-producing E. coli can induce immunogenic cell death in tumor tissues, potentially acting as adjuvants to immunotherapy [1111Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.]. However, this immune activation can also trigger excessive inflammation or autoimmunity. For instance, lipopolysaccharide (LPS) on E. coli surfaces can provoke Toll-like receptor (TLR)-mediated cytokine storms [99Zhao J, Liu H, Zhang L, Yang C. Hypoxia-responsive bacterial therapy for glioblastoma. Nat Commun. 2023;14:1723.]. Fine-tuning the immunostimulatory profile, such as by LPS modification or strain reduction, is critical for maximizing therapeutic effect while minimizing risk [3030Cai S, Li J, Zhao H. Programmable kill-switch systems for biosafety in engineered therapeutic bacteria. ACS Synth Biol. 2022;11(4):1342–1354.].
Manufacturing, scalability and delivery
Unlike conventional drugs, engineered E. coli must be cultured, preserved, and delivered without losing function. Maintaining plasmid stability, viability, and biosafety features during large-scale manufacturing poses logistical hurdles [2929Gao X, Park JH, Xu H. Auxotrophy-based biosafety systems in engineered microbes. Biotechnol Adv. 2021;49:107757.]. Freeze-drying (lyophilization), encapsulation in protective hydrogels, or storage under anaerobic conditions are being tested [3232Huang P, Shen X, Wang L. Hydrogel-encapsulated engineered bacteria for sustained intracranial therapy in glioma models. ACS Biomater Sci Eng. 2024;10(6):2134–2145.]. Moreover, E. coli transport to intracranial tumors via the Blood-brain Barrier (BBB) is ineffective. Focused Ultrasound (FUS)-assisted BBB opening, hydrogel microsphere delivery, and lipid nanocarriers are also being investigated to increase targeted delivery and retention [3131Turner L, Houghton J, Danino T. Hydrogel-based encapsulation of engineered bacteria for localized glioblastoma therapy. Biomaterials. 2023;292:121932.].
Future perspectives
To overcome the constraints preventing clinical translation of modified Escherichia coli (E. coli) therapeutics for glioblastoma, future research should target five crucial domains:
Clinical-grade biocontainment: Ensuring patient safety is critical when delivering live bacterial treatments. Advanced biocontainment systems that include numerous safety layers, such as auxotrophy, quorum-sensing death switches, and CRISPR-based genetic confinement, must be adjusted for evolutionary stability and performance in vivo [4949Hayashi T, Suda W, Shibata N. Multi-layered biocontainment strategies for live bacterial therapeutics. Nat Biotechnol. 2024;42(4):675–688.]. These protections will reduce the hazards of bacterial escape, systemic infection, and horizontal gene transfer.
Logic-gated payload control: Next-generation therapeutic strains should be equipped with logic-gated genetic circuits that trigger ROS generation or drug release only in response to specific tumor microenvironmental cues, such as hypoxia, lactate accumulation, or acidic pH. This dual- or multi-signal gating improves selectivity and reduces off-target effects [1616Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.].
Combination therapeutic approaches: The integration of engineered E. coli therapies with existing modalities, such as immune checkpoint inhibitors, radiotherapy, and temozolomide, offers a synergistic strategy to overcome tumor resistance and enhance antitumor immunity. Such combinatorial approaches can exploit ROS-induced immunogenic cell death to prime the tumor for immune engagement [1111Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.].
Advanced preclinical models: Translational fidelity remains a major gap. Current rodent models inadequately mimic human GBM heterogeneity and immune complexity. Therefore, humanized animal models, 3D tumor organoids, and patient-derived xenograft (PDX) systems are essential to assess efficacy, immunotoxicity, and spatially variable responses [2828Huang R, Xu Y, Chen Y, Zhao L. Smart nanogels for targeted delivery of bacterial therapeutics in glioblastoma. Adv Drug Deliv Rev. 2024;200:115104.].
Regulatory collaboration: Establishing clear regulatory frameworks tailored to live microbial therapeutics is essential. Ongoing dialogue between researchers, regulatory bodies, and bioethicists will be required to develop approval pathways that address unique concerns associated with genetically modified organisms (GMOs), including biocontainment, long-term monitoring, and environmental impact [3333Singh A, Gupta S, Sharma R. Stimuli-responsive nanogels for bacterial drug delivery in glioblastoma treatment. J Control Release. 2022;350:234–246.].
By investing in these strategic areas, the field can advance from proof-of-concept to viable clinical applications, potentially offering transformative therapies for patients with GBM.
Engineered Escherichia coli strains represent a groundbreaking approach in glioblastoma therapy, offering precision-targeted delivery of reactive oxygen species (ROS) for selective tumor destruction and immune activation. Preclinical studies have shown encouraging efficacy, especially when combined with immunotherapy or advanced delivery systems. However, significant translational barriers remain—including biosafety, tumor heterogeneity, immune compatibility, and regulatory hurdles. Addressing these challenges through robust biocontainment strategies, adaptive genetic circuits, advanced preclinical models, and collaborative regulatory frameworks is essential. With continued innovation and validation, E. coli-based ROS therapeutics may soon emerge as a clinically viable, next-generation treatment for glioblastoma.
Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.
Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, Gorlia T, Allgeier A, Lacombe D, Cairncross JG, Eisenhauer E, Mirimanoff RO; European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups; National Cancer Institute of Canada Clinical Trials Group. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005 Mar 10;352(10):987-96. doi: 10.1056/NEJMoa043330. PMID: 15758009.
Ostrom QT, Price M, Neff C, Cioffi G, Waite KA, Kruchko C, Barnholtz-Sloan JS. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2015-2019. Neuro Oncol. 2022 Oct 5;24(Suppl 5):v1-v95. doi: 10.1093/neuonc/noac202. PMID: 36196752; PMCID: PMC9533228.
Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 2006 Jan;7(1):41-53. doi: 10.1038/nrn1824. PMID: 16371949.
Guan X, Wang C, Zhang L, Zhang Y. Reactive oxygen species in glioblastoma: From pathophysiology to therapeutic strategy. Front Oncol. 2023;13:1184627.
Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020 Aug 10;38(2):167-197. doi: 10.1016/j.ccell.2020.06.001. Epub 2020 Jul 9. PMID: 32649885; PMCID: PMC7439808.
Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.
Zhao J, Liu H, Zhang L, Yang C. Hypoxia-responsive bacterial therapy for glioblastoma. Nat Commun. 2023;14:1723.
Chen Y, Liu X, Zhang C, Zhao T, Xu H, Li F. Synthetic biology-engineered Escherichia coli for targeted glioblastoma therapy via ROS-mediated mechanisms. Nat Biomed Eng. 2022;6(9):1035–1048.
Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.
Guan Y, Wang C, Luo L. ROS-mediated therapies in glioblastoma: Molecular targets and delivery strategies. Cell Mol Life Sci. 2023;80:56.
Chen Y, Wang H, Xu Y, Li R. Engineered Escherichia coli for tumor-targeted prodrug therapy in glioblastoma. Nat Biomed Eng. 2022;6(12):1407–1419.
Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.
Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.
Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.
Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.
Li F, Zhang H, Yang M. Engineered bacterial carriers for ROS-based synergistic glioblastoma therapy and immune modulation. Adv Drug Deliv Rev. 2024;199:114982.
Quail DF, Joyce JA. The Microenvironmental Landscape of Brain Tumors. Cancer Cell. 2017 Mar 13;31(3):326-341. doi: 10.1016/j.ccell.2017.02.009. PMID: 28292436; PMCID: PMC5424263.
Meng F, Evans CW, Bhati M, Danino T. Hypoxia-inducible bacterial therapies for precision oncology. Nat Biomed Eng. 2022;6(8):932–946.
Kroemer G, Galassi C, Zitvogel L, Galluzzi L. Immunogenic cell death in cancer therapy. Annu Rev Immunol. 2022;40:17–36.
Choi J, Yang H, Kim E, Kim J. Synthetic biology approaches to reprogramming immune responses in cancer therapy. Nat Rev Immunol. 2022;22(10):618–635.
Xu H, Zhang P, Huang S. Synergistic glioblastoma treatment with engineered E. coli and immune checkpoint inhibitors. Sci Adv. 2023;9(12):eadg3457.
Zhou S, Gravekamp C, Bermudes D, Liu K. Tumour-targeting bacteria engineered to fight cancer. Nat Rev Cancer. 2021;21(11):709–722.
Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.
Turner JD, Miller CV, Gibson L. Thermo-responsive PEG–PCL hydrogels for intracranial bacterial therapy. Biomaterials. 2023;294:121027.
Huang R, Xu Y, Chen Y, Zhao L. Smart nanogels for targeted delivery of bacterial therapeutics in glioblastoma. Adv Drug Deliv Rev. 2024;200:115104.
Gao X, Park JH, Xu H. Auxotrophy-based biosafety systems in engineered microbes. Biotechnol Adv. 2021;49:107757.
Cai S, Li J, Zhao H. Programmable kill-switch systems for biosafety in engineered therapeutic bacteria. ACS Synth Biol. 2022;11(4):1342–1354.
Turner L, Houghton J, Danino T. Hydrogel-based encapsulation of engineered bacteria for localized glioblastoma therapy. Biomaterials. 2023;292:121932.
Huang P, Shen X, Wang L. Hydrogel-encapsulated engineered bacteria for sustained intracranial therapy in glioma models. ACS Biomater Sci Eng. 2024;10(6):2134–2145.
Singh A, Gupta S, Sharma R. Stimuli-responsive nanogels for bacterial drug delivery in glioblastoma treatment. J Control Release. 2022;350:234–246.
Shah N, Miura Y, Kohane DS. Transferrin-modified liposomal carriers enhance rodent glioma drug delivery. Nanomedicine. 2019;14(18):2317–2330.
Kong L, Zhou Q, Cao H. Dendrimer-entrapped gold nanoparticles functionalized with RGD enhance siRNA delivery across the BBB. ACS Nano. 2016;10(5):5530–5541.
Singh R, Malhotra R, Rawat P. Regulatory landscape of live microbial therapeutics. J Biomed Regul. 2022;15(2):88–97.
Luo D, Pan Y, Zhang X. Angiopep-2 peptide-labeled RBC nanocarrier for multimodal BBB crossing and GBM therapy. J Nanobiotechnology. 2021;19(1):17.
Liang C, Zhang Y, Zhou J. Macrophage-membrane-coated nanoparticles for BBB penetration and GBM therapy. Adv Funct Mater. 2023;33(12):2300650.
Ma X, Wang S, Li D. Biomimetic glioma-cell-coated nanocarriers for targeted chemotherapy delivery. J Nanobiotechnology. 2024;22(1):12.
Dong L, Zhao R, Wu J. RBC membrane–coated bacterial vesicles improve GBM target delivery and survival in mice. Biomaterials. 2023;351:121093.
Meng Y, Hynynen K, Lipsman N. Focused ultrasound in the brain: From thermoablation to drug delivery. Nat Rev Neurol. 2018;14(10):585–599.
Rogers EM, Patel RB, Smith CM. Focused ultrasound–mediated delivery of engineered bacteria to brain tumors. Theranostics. 2022;12(9):3667–3680.
Ahangarzadeh S, Rezaei R, Mahjoubin-Tehran M, Pourhanifeh MH, Rajabibazl M, Hamblin MR. Outer membrane vesicles as a new platform in glioblastoma therapy. Front Oncol. 2023;13:1119256.
Rogers W, Han X, Chen Y. Focused ultrasound-mediated blood–brain barrier modulation for bacterial therapy in glioblastoma. Front Bioeng Biotechnol. 2022;10:942865.
Singh R, Malhotra R, Rawat P. Targeted nanogel systems for BBB transcytosis and glioblastoma drug delivery. J Biomed Nanotechnol. 2022;18(1):104–120.
Shah B, Mishra V, Kesharwani P, Jain NK. Outer membrane vesicles: New insights into delivery platforms for glioblastoma therapy. Mol Pharm. 2019;16(9):3787–3798.
Ng WL, Bassler BL. Bacterial quorum-sensing network architectures. Annu Rev Genet. 2009;43:197-222. doi: 10.1146/annurev-genet-102108-134304. PMID: 19686078; PMCID: PMC4313539.
Gurbatri CR, Lia I, Vincent R, Coker C, Castro S, Treuting PM, Hinchliffe TE, Arpaia N, Danino T. Engineered probiotics for local tumor delivery of checkpoint blockade nanobodies. Sci Transl Med. 2020 Feb 12;12(530):eaax0876. doi: 10.1126/scitranslmed.aax0876. PMID: 32051224; PMCID: PMC7685004.
Hayashi T, Suda W, Shibata N. Multi-layered biocontainment strategies for live bacterial therapeutics. Nat Biotechnol. 2024;42(4):675–688.
Prinz M, Masuda T, Wheeler MA, Quintana FJ. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu Rev Immunol. 2021 Apr 26;39:251-277. doi: 10.1146/annurev-immunol-093019-110159. Epub 2021 Feb 8. PMID: 33556248; PMCID: PMC8085109.
Weller M, Wick W, Aldape K, Brada M, Berger M, Pfister SM, Nishikawa R, Rosenthal M, Wen PY, Stupp R, Reifenberger G. Glioma. Nat Rev Dis Primers. 2015 Jul 16;1:15017. doi: 10.1038/nrdp.2015.17. PMID: 27188790.
Roberts NJ, Zhang L, Janku F, Collins A, Bai RY, Staedtke V, Rusk AW, Tung D, Miller M, Roix J, Khanna KV, Murthy R, Benjamin RS, Helgason T, Szvalb AD, Bird JE, Roy-Chowdhuri S, Zhang HH, Qiao Y, Karim B, McDaniel J, Elpiner A, Sahora A, Lachowicz J, Phillips B, Turner A, Klein MK, Post G, Diaz LA Jr, Riggins GJ, Papadopoulos N, Kinzler KW, Vogelstein B, Bettegowda C, Huso DL, Varterasian M, Saha S, Zhou S. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci Transl Med. 2014 Aug 13;6(249):249ra111. doi: 10.1126/scitranslmed.3008982. PMID: 25122639; PMCID: PMC4399712.
Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.
Mandell DJ, Lajoie MJ, Mee MT, Takeuchi R, Kuznetsov G, Norville JE, Gregg CJ, Stoddard BL, Church GM. Biocontainment of genetically modified organisms by synthetic protein design. Nature. 2015 Feb 5;518(7537):55-60. doi: 10.1038/nature14121. Epub 2015 Jan 21. Erratum in: Nature. 2015 Nov 12;527(7577):264. doi: 10.1038/nature15536. PMID: 25607366; PMCID: PMC4422498.
Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.
Godonu SS. Engineered Escherichia coli Strains as Therapeutic Agents in Reactive Oxygen Species (ROS)-Mediated Glioblastoma Treatment: A Systematic Review of Mechanisms, Efficacy and Challenges. IgMin Res. August 19, 2025; 3(8): 285-295. IgMin ID: igmin310; DOI:10.61927/igmin310; Available at: igmin.link/p310
Address Correspondence: Sewanu Stephen Godonu, American Institute of Medical Sciences, Jamaica, Email: [email protected]
How to cite this article: Godonu SS. Engineered Escherichia coli Strains as Therapeutic Agents in Reactive Oxygen Species (ROS)-Mediated Glioblastoma Treatment: A Systematic Review of Mechanisms, Efficacy and Challenges. IgMin Res. August 19, 2025; 3(8): 285-295. IgMin ID: igmin310; DOI:10.61927/igmin310; Available at: igmin.link/p310
Figure 1: Risks of Endotoxemia and Mitigation strategies in ...
Table 1: Summary of representative preclinical studies usin...
Table 2: Key delivery and targeting platforms for engineere...
Table 3: Matrix summarizing major risks arising from engine...
Lim M, Xia Y, Bettegowda C, Weller M. Current state of immunotherapy for glioblastoma. Nat Rev Clin Oncol. 2018 Jul;15(7):422-442. doi: 10.1038/s41571-018-0003-5. PMID: 29643471.
Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, Gorlia T, Allgeier A, Lacombe D, Cairncross JG, Eisenhauer E, Mirimanoff RO; European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups; National Cancer Institute of Canada Clinical Trials Group. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005 Mar 10;352(10):987-96. doi: 10.1056/NEJMoa043330. PMID: 15758009.
Ostrom QT, Price M, Neff C, Cioffi G, Waite KA, Kruchko C, Barnholtz-Sloan JS. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2015-2019. Neuro Oncol. 2022 Oct 5;24(Suppl 5):v1-v95. doi: 10.1093/neuonc/noac202. PMID: 36196752; PMCID: PMC9533228.
Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 2006 Jan;7(1):41-53. doi: 10.1038/nrn1824. PMID: 16371949.
Guan X, Wang C, Zhang L, Zhang Y. Reactive oxygen species in glioblastoma: From pathophysiology to therapeutic strategy. Front Oncol. 2023;13:1184627.
Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020 Aug 10;38(2):167-197. doi: 10.1016/j.ccell.2020.06.001. Epub 2020 Jul 9. PMID: 32649885; PMCID: PMC7439808.
Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring LS, Bhatia SN, Hasty J. Synchronized cycles of bacterial lysis for in vivo delivery. Nature. 2016 Aug 4;536(7614):81-85. doi: 10.1038/nature18930. Epub 2016 Jul 20. PMID: 27437587; PMCID: PMC5048415.
Zhao J, Liu H, Zhang L, Yang C. Hypoxia-responsive bacterial therapy for glioblastoma. Nat Commun. 2023;14:1723.
Chen Y, Liu X, Zhang C, Zhao T, Xu H, Li F. Synthetic biology-engineered Escherichia coli for targeted glioblastoma therapy via ROS-mediated mechanisms. Nat Biomed Eng. 2022;6(9):1035–1048.
Wang H, Yu C, Gao X, Guo L. Oxidative stress-mediated glioblastoma therapy: Advances and challenges. Cell Death Dis. 2022;13:533.
Guan Y, Wang C, Luo L. ROS-mediated therapies in glioblastoma: Molecular targets and delivery strategies. Cell Mol Life Sci. 2023;80:56.
Chen Y, Wang H, Xu Y, Li R. Engineered Escherichia coli for tumor-targeted prodrug therapy in glioblastoma. Nat Biomed Eng. 2022;6(12):1407–1419.
Wang Z, Liu Q, Ren J. Programmable probiotic therapy with ROS-inducing bacteria for glioblastoma immunotherapy. ACS Nano. 2022;16(12):19132–19145.
Li Y, Zhang J, Wang X, Sun Q. Engineered microbial systems for redox-based cancer therapy. Adv Drug Deliv Rev. 2024;199:115071.
Xu Y, Li R, Chen Y, Zhao L. Engineered bacteria synergize with immune checkpoint inhibitors for glioblastoma treatment. Nat Commun. 2023;14:6459.
Zhao T, Liu B, Sun W. Bioengineered E. coli for ROS delivery: Tumor specificity via hypoxia promoters and quorum circuits. J Control Release. 2023;356:572–583.
Li F, Zhang H, Yang M. Engineered bacterial carriers for ROS-based synergistic glioblastoma therapy and immune modulation. Adv Drug Deliv Rev. 2024;199:114982.
Quail DF, Joyce JA. The Microenvironmental Landscape of Brain Tumors. Cancer Cell. 2017 Mar 13;31(3):326-341. doi: 10.1016/j.ccell.2017.02.009. PMID: 28292436; PMCID: PMC5424263.
Meng F, Evans CW, Bhati M, Danino T. Hypoxia-inducible bacterial therapies for precision oncology. Nat Biomed Eng. 2022;6(8):932–946.
Kroemer G, Galassi C, Zitvogel L, Galluzzi L. Immunogenic cell death in cancer therapy. Annu Rev Immunol. 2022;40:17–36.
Choi J, Yang H, Kim E, Kim J. Synthetic biology approaches to reprogramming immune responses in cancer therapy. Nat Rev Immunol. 2022;22(10):618–635.
Xu H, Zhang P, Huang S. Synergistic glioblastoma treatment with engineered E. coli and immune checkpoint inhibitors. Sci Adv. 2023;9(12):eadg3457.
Zhou S, Gravekamp C, Bermudes D, Liu K. Tumour-targeting bacteria engineered to fight cancer. Nat Rev Cancer. 2021;21(11):709–722.
Riglar DT, Silver PA. Engineering bacteria for diagnostic and therapeutic applications. Nat Rev Microbiol. 2018 Apr;16(4):214-225. doi: 10.1038/nrmicro.2017.172. Epub 2018 Feb 5. PMID: 29398705.
Turner JD, Miller CV, Gibson L. Thermo-responsive PEG–PCL hydrogels for intracranial bacterial therapy. Biomaterials. 2023;294:121027.
Huang R, Xu Y, Chen Y, Zhao L. Smart nanogels for targeted delivery of bacterial therapeutics in glioblastoma. Adv Drug Deliv Rev. 2024;200:115104.
Gao X, Park JH, Xu H. Auxotrophy-based biosafety systems in engineered microbes. Biotechnol Adv. 2021;49:107757.
Cai S, Li J, Zhao H. Programmable kill-switch systems for biosafety in engineered therapeutic bacteria. ACS Synth Biol. 2022;11(4):1342–1354.
Turner L, Houghton J, Danino T. Hydrogel-based encapsulation of engineered bacteria for localized glioblastoma therapy. Biomaterials. 2023;292:121932.
Huang P, Shen X, Wang L. Hydrogel-encapsulated engineered bacteria for sustained intracranial therapy in glioma models. ACS Biomater Sci Eng. 2024;10(6):2134–2145.
Singh A, Gupta S, Sharma R. Stimuli-responsive nanogels for bacterial drug delivery in glioblastoma treatment. J Control Release. 2022;350:234–246.
Shah N, Miura Y, Kohane DS. Transferrin-modified liposomal carriers enhance rodent glioma drug delivery. Nanomedicine. 2019;14(18):2317–2330.
Kong L, Zhou Q, Cao H. Dendrimer-entrapped gold nanoparticles functionalized with RGD enhance siRNA delivery across the BBB. ACS Nano. 2016;10(5):5530–5541.
Singh R, Malhotra R, Rawat P. Regulatory landscape of live microbial therapeutics. J Biomed Regul. 2022;15(2):88–97.
Luo D, Pan Y, Zhang X. Angiopep-2 peptide-labeled RBC nanocarrier for multimodal BBB crossing and GBM therapy. J Nanobiotechnology. 2021;19(1):17.
Liang C, Zhang Y, Zhou J. Macrophage-membrane-coated nanoparticles for BBB penetration and GBM therapy. Adv Funct Mater. 2023;33(12):2300650.
Ma X, Wang S, Li D. Biomimetic glioma-cell-coated nanocarriers for targeted chemotherapy delivery. J Nanobiotechnology. 2024;22(1):12.
Dong L, Zhao R, Wu J. RBC membrane–coated bacterial vesicles improve GBM target delivery and survival in mice. Biomaterials. 2023;351:121093.
Meng Y, Hynynen K, Lipsman N. Focused ultrasound in the brain: From thermoablation to drug delivery. Nat Rev Neurol. 2018;14(10):585–599.
Rogers EM, Patel RB, Smith CM. Focused ultrasound–mediated delivery of engineered bacteria to brain tumors. Theranostics. 2022;12(9):3667–3680.
Ahangarzadeh S, Rezaei R, Mahjoubin-Tehran M, Pourhanifeh MH, Rajabibazl M, Hamblin MR. Outer membrane vesicles as a new platform in glioblastoma therapy. Front Oncol. 2023;13:1119256.
Rogers W, Han X, Chen Y. Focused ultrasound-mediated blood–brain barrier modulation for bacterial therapy in glioblastoma. Front Bioeng Biotechnol. 2022;10:942865.
Singh R, Malhotra R, Rawat P. Targeted nanogel systems for BBB transcytosis and glioblastoma drug delivery. J Biomed Nanotechnol. 2022;18(1):104–120.
Shah B, Mishra V, Kesharwani P, Jain NK. Outer membrane vesicles: New insights into delivery platforms for glioblastoma therapy. Mol Pharm. 2019;16(9):3787–3798.
Ng WL, Bassler BL. Bacterial quorum-sensing network architectures. Annu Rev Genet. 2009;43:197-222. doi: 10.1146/annurev-genet-102108-134304. PMID: 19686078; PMCID: PMC4313539.
Gurbatri CR, Lia I, Vincent R, Coker C, Castro S, Treuting PM, Hinchliffe TE, Arpaia N, Danino T. Engineered probiotics for local tumor delivery of checkpoint blockade nanobodies. Sci Transl Med. 2020 Feb 12;12(530):eaax0876. doi: 10.1126/scitranslmed.aax0876. PMID: 32051224; PMCID: PMC7685004.
Hayashi T, Suda W, Shibata N. Multi-layered biocontainment strategies for live bacterial therapeutics. Nat Biotechnol. 2024;42(4):675–688.
Prinz M, Masuda T, Wheeler MA, Quintana FJ. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu Rev Immunol. 2021 Apr 26;39:251-277. doi: 10.1146/annurev-immunol-093019-110159. Epub 2021 Feb 8. PMID: 33556248; PMCID: PMC8085109.
Weller M, Wick W, Aldape K, Brada M, Berger M, Pfister SM, Nishikawa R, Rosenthal M, Wen PY, Stupp R, Reifenberger G. Glioma. Nat Rev Dis Primers. 2015 Jul 16;1:15017. doi: 10.1038/nrdp.2015.17. PMID: 27188790.
Roberts NJ, Zhang L, Janku F, Collins A, Bai RY, Staedtke V, Rusk AW, Tung D, Miller M, Roix J, Khanna KV, Murthy R, Benjamin RS, Helgason T, Szvalb AD, Bird JE, Roy-Chowdhuri S, Zhang HH, Qiao Y, Karim B, McDaniel J, Elpiner A, Sahora A, Lachowicz J, Phillips B, Turner A, Klein MK, Post G, Diaz LA Jr, Riggins GJ, Papadopoulos N, Kinzler KW, Vogelstein B, Bettegowda C, Huso DL, Varterasian M, Saha S, Zhou S. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci Transl Med. 2014 Aug 13;6(249):249ra111. doi: 10.1126/scitranslmed.3008982. PMID: 25122639; PMCID: PMC4399712.
Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.
Mandell DJ, Lajoie MJ, Mee MT, Takeuchi R, Kuznetsov G, Norville JE, Gregg CJ, Stoddard BL, Church GM. Biocontainment of genetically modified organisms by synthetic protein design. Nature. 2015 Feb 5;518(7537):55-60. doi: 10.1038/nature14121. Epub 2015 Jan 21. Erratum in: Nature. 2015 Nov 12;527(7577):264. doi: 10.1038/nature15536. PMID: 25607366; PMCID: PMC4422498.
Chan CTY, Lee JW, Cameron DE, Bashor CJ, Collins JJ. ‘Deadman’ and ‘Passcode’ microbial kill switches for bacterial containment. Nat Chem Biol. 2016;12(2):82–86.
Engineered Escherichia coli Strains as Therapeutic Agents in Reactive Oxygen Species (ROS)-Mediated Glioblastoma Treatment: A Systematic Review of Mechanisms, Efficacy and Challenges



 Scan and get link
Scan and get link