IGMIN: We're glad you're here. Please click 'create a new query' if you are a new visitor to our website and need further information from us.
If you are already a member of our network and need to keep track of any developments regarding a question you have already submitted, click 'take me to my Query.'
Welcome to IgMin Research – an Open Access journal uniting Biology, Medicine, and Engineering. We’re dedicated to advancing global knowledge and fostering collaboration across scientific fields.
At IgMin Research, we bridge the frontiers of Biology, Medicine, and Engineering to foster interdisciplinary innovation. Our expanded scope now embraces a wide spectrum of scientific disciplines, empowering global researchers to explore, contribute, and collaborate through open access.
Welcome to IgMin, a leading platform dedicated to enhancing knowledge dissemination and professional growth across multiple fields of science, technology, and the humanities. We believe in the power of open access, collaboration, and innovation. Our goal is to provide individuals and organizations with the tools they need to succeed in the global knowledge economy.
IgMin Publications Inc., Suite 102, West Hartford, CT - 06110, USA
Medicine GroupResearch ArticleArticle ID: igmin114
The Antioxidant and Antidepressant Properties of Dietary Proteins Derived from Egg and Bean Extracts and Their Acute Toxicity: A Journey from Nutrition to Pharmacognosy
This research reveals the previously unexplored pharmacognostic potential of antidepressants found in nutrients derived from both legume and animal sources. Through preclinical investigations involving mouse models, the study focused into antidepressant and antioxidant activities of non-denatured and denatured protein extracts from beans and eggs. Non-denatured protein extracts from beans and eggs, at saturation levels of 40% and 80%, were examined as macronutrients, while denatured protein extracts at equivalent saturation levels were considered micronutrients. The study employed the DPPH and hydrogen peroxide tests to assess antioxidant activity, and the forced swimming test and sucrose preference test to evaluate acute and chronic mild antidepressant effects, respectively. The acute toxicity study revealed that macronutrients from eggs at 40% and 80% saturation displayed non-toxic effects (LD50 >5 g/kg), while those from beans, specifically at saturation of 80%, exhibited a relatively low level of toxicity (LD50 = 2.5 g/kg). Evaluation of antioxidant activity using the DPPH test yielded inconclusive results due to the influence of ethanol precipitation. In contrast, the H2O2 test demonstrated significant antioxidant potential in both macronutrients and micronutrients extracted from beans and eggs at all saturation levels. In investigating antidepressant properties, both macronutrients and micronutrients of bean and egg protein extracts at 40% and 80% saturation exhibited notable antidepressant effects, particularly the micronutrients at saturation of 80%. This antidepressant effect was characterized by a reduction in immobility time and an increase in sucrose preference.
In conclusion, this study uncovers the multifaceted potential of protein extracts sourced from natural products, plant and animal origins, as agents for treating depression. It opens up new avenues for research, with implications ranging from neuroprotection to the management of depression, inspiring optimism for innovative approaches to mental health treatment.
In recent years, the intricate connection between diet and mental health has garnered increasing attention within the realm of scientific inquiry. Among the various dietary components, proteins have emerged as crucial players not only in maintaining physical health but also in influencing mental well-being [11Smith K. The intricate relationship between diet, mental health, and well-being. Frontiers in Human Neuroscience. 2019; 13(33).-33Gómez-Pinilla F. Brain foods: the effects of nutrients on brain function. Nat Rev Neurosci. 2008 Jul;9(7):568-78. doi: 10.1038/nrn2421. PMID: 18568016; PMCID: PMC2805706.]. The impact of dietary proteins on the complex landscape of depression has become a subject of considerable interest, as researchers seek to uncover the potential benefits they may offer in mitigating depressive symptoms [11Smith K. The intricate relationship between diet, mental health, and well-being. Frontiers in Human Neuroscience. 2019; 13(33).,22Sarris J, Logan AC, Akbaraly TN, Amminger GP, Balanzá-Martínez V, Freeman MP, Hibbeln J, Matsuoka Y, Mischoulon D, Mizoue T, Nanri A, Nishi D, Ramsey D, Rucklidge JJ, Sanchez-Villegas A, Scholey A, Su KP, Jacka FN; International Society for Nutritional Psychiatry Research. Nutritional medicine as mainstream in psychiatry. Lancet Psychiatry. 2015 Mar;2(3):271-4. doi: 10.1016/S2215-0366(14)00051-0. Epub 2015 Feb 25. PMID: 26359904.].
Depression, a prevalent and debilitating mental disorder, poses a significant global health burden. Its multifaceted etiology involves a combination of genetic, environmental, and neurochemical factors [44Belmaker RH, Agam G. Major depressive disorder. N Engl J Med. 2008 Jan 3;358(1):55-68. doi: 10.1056/NEJMra073096. PMID: 18172175.]. Traditional treatment approaches often involve psychotherapy and pharmacotherapy, yet the quest for innovative and holistic interventions has led researchers to explore the potential influence of diet on mental health outcomes []. In this context, proteins, the essential building blocks of life, have emerged as not only nutritional necessities but also as agents that might impact the brain’s chemistry and functioning [66Lieberman MD, Gaunt R, Gilbert DT, Trope Y. Reflexion and reflection: A social cognitive neuroscience approach to attributional inference. Advances in experimental social psychology. 2002; 34:199‑].
Proteins are instrumental in a multitude of physiological processes, including the synthesis of neurotransmitters, enzymes, and hormones [77Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell. 4th Garland Science; 2002.]. These bioactive compounds play a pivotal role in regulating mood, cognition, and emotional well-being [88Fernstrom JD. Large neutral amino acids: dietary effects on brain neurochemistry and function. Amino Acids. 2013 Sep;45(3):419-30. doi: 10.1007/s00726-012-1330-y. Epub 2012 Jun 8. PMID: 22677921.]. Recent studies have illuminated the intricate relationship between dietary protein intake and the intricate neural pathways that govern depression [22Sarris J, Logan AC, Akbaraly TN, Amminger GP, Balanzá-Martínez V, Freeman MP, Hibbeln J, Matsuoka Y, Mischoulon D, Mizoue T, Nanri A, Nishi D, Ramsey D, Rucklidge JJ, Sanchez-Villegas A, Scholey A, Su KP, Jacka FN; International Society for Nutritional Psychiatry Research. Nutritional medicine as mainstream in psychiatry. Lancet Psychiatry. 2015 Mar;2(3):271-4. doi: 10.1016/S2215-0366(14)00051-0. Epub 2015 Feb 25. PMID: 26359904.]. Amino acids, the foundational components of proteins, are essential precursors to neurotransmitters such as serotonin and dopamine, which are closely linked to mood regulation [99Fernstrom JD, Fernstrom MH. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J Nutr. 2007 Jun;137(6 Suppl 1):1539S-1547S; discussion 1548S. doi: 10.1093/jn/137.6.1539S. PMID: 17513421.].
With the objective of achieving this aim, the current study was undertaken to investigate the antidepressant and antioxidant attributes of protein extracts sourced from beans and eggs. These extracts underwent analysis at saturation levels of 40% and 80%, with categorization as macronutrients in their undenatured state, and as micronutrients when subjected to denaturation.
Food materials: The food materials used consist of eggs and beans. The materials were purchased from a grocery store.
Animal materials:In vivo experiments were carried out on both male and female mice. These mice have a weight ranging from 120g to 360g, and they have unrestricted access to water and food.
Methods
Preparation of protein extracts: Firstly, a bean macerate was prepared by adding 10g of bean powder to 100 mL of physiological solution (0.9% NaCl, pH 7.4). Then, the extracts were allowed to macerate for 24 hours at 4 °C. After maceration, the mixture was centrifuged (for 15 minutes at 2700 g at 4 °C) and the resulting supernatant was sterilized through a 0.45 µm nitrocellulose filter. In parallel, an egg extract was prepared by diluting egg white at a 1/12 ratio. Subsequently, the bean and egg protein extracts were prepared by adding ammonium sulfate at concentrations of 40% and 80% (w/v) at 4 °C [1010Englard S, Seifter S. Precipitation techniques. Methods Enzymol. 1990;182:285-300. doi: 10.1016/0076-6879(90)82024-v. PMID: 2314242.]. Then, the samples were subsequently stored until needed.
Heat treatment: This process entails denaturing the proteins found in the protein extracts of eggs and beans. To accomplish this, protein extracts used to be heated for 30 minutes in a water bath maintained at 100 °C. In order to prevent any evaporation of sample water, the containers were hermetically sealed. After the treatment, the samples were cooled in a water bath maintained at 25 °C and then left at room temperature for about an hour before being stored in the cold room at 4 °C [1111Boye JI, Aksay S, Roufik S, Ribereau S, Mondor M, Farnworth E, Rajamohamed SH. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Research International. 2010; 43:537‑].
Acute toxicity study: We examined the acute toxicity of the egg and bean extracts in mice weighing between 120 and 360g. The mice were organized into seven groups of four each (n = 4; 2 male and 2 female). After administering the extracts intraperitoneally (i.p), ranging from 0.7 to 5g/Kg body weight (b.w.), we observed and recorded their overall behavior. At the culmination of the study, the animals were administered sodium pentobarbital for anesthesia, followed by euthanasia. Essential organs, including the liver, kidneys, spleen, lungs, and heart, were carefully removed, rinsed with a solution of 0.9% NaCl, and then weighed. The relative weight of each organ was determined using the subsequent formula [1212Reduan FH, Shaari RM, Sayuti NSA, Mustapha NM, Abu Bakar MZ, Sithambaram S, Hamzah H. Acute and subacute dermal toxicity of ethanolic extract of Melastoma malabathricumleaves in Sprague-Dawley rats. Toxicol Res. 2020 Mar 26;36(3):203-210. doi: 10.1007/s43188-019-00013-5. PMID: 32685424; PMCID: PMC7352010.]:
RW= (W0/BW) ×100
Where:
RW: Relative weight of the organ.
W0: Weight of the organ in grams.
BW: Body weight of the rat in grams.
Antioxidant assays
- DPPH assay
We evaluated the antioxidant potential of the examined extracts through the employment of the 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay as outlined in the study by Qamar, et al. [1313Qamar F,Naveed S, Faizi S, Sana A. Formulation and Evaluation of Natural Antioxidant Cream of Ocimum basilicum. Latin American Journal of Pharmacy. 2021; 40:2293‑]. According to this procedure, various concentrations (5-200 µg/mL) of the tested extracts and ascorbic acid (used as a reference) were prepared. Subsequently, 1mL of each tested extract was combined with 3mL of pre-dissolved DPPH (0.004%) in ethanol. The mixture was thoroughly mixed and then allowed to incubate for 30 minutes at room temperature under light protection. Following this incubation period, the spectrophotometric measurement, using a Zuzi spectrophotometer, of the solutions’ absorbance at 517 nm against a blank containing only ethanol was conducted. The inhibition percentage (I%) of the DPPH free radical was then calculated using the formula:
I% = [(Ablank-Asample)/Ablank] × 100
- Hydrogen peroxide assay
The antioxidant potential of the extracts using the H2O2 test was evaluated as described by Qamar, et al. [1313Qamar F,Naveed S, Faizi S, Sana A. Formulation and Evaluation of Natural Antioxidant Cream of Ocimum basilicum. Latin American Journal of Pharmacy. 2021; 40:2293‑]. A series of concentrations for the studied protein extracts (5-200 µg/mL) and ascorbic acid were prepared beforehand. Next, 1mL of the tested extracts was combined with 0.6mL of H2O2 (40 mM). Following this, the reaction mixture was incubated at room temperature for 10 minutes. Afterward, the absorbance of H2O2 was determined using a UV-Vis spectrophotometer (model 6300, VWR) at a wavelength of 230 nm. The percentage of H2O2 inhibition was calculated using the subsequent formula:
I (%) = [(Ablank-Asample)/Ablank] x 100
Forced swim test: This test was employed to investigate the antidepressant effects of protein extracts from eggs and beans on acute depression [1414Krishnan V, Nestler EJ. Animal models of depression: molecular perspectives. Curr Top Behav Neurosci. 2011;7:121-47. doi: 10.1007/7854_2010_108. PMID: 21225412; PMCID: PMC3270071.]. The forced swim test, a recognized behavioral assessment tool, was utilized as described by Porsolt, et al. [1515Porsolt RD, Le Pichon M, Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977 Apr 21;266(5604):730-2. doi: 10.1038/266730a0. PMID: 559941.]. Prior to the test, the mice were divided into twenty groups, with each group consisting of four animals (n = 4; 2 females and 2 males). Subsequently, the protein extracts were administered via “i.p” at various doses ranging from 14 µg/Kg (b.w.) to 140 µg/Kg (b.w.) for the test groups. The control groups were received a solution of 0.9% NaCl at a pH of 7.4, while the reference groups were treated with sertraline, a reference molecule known for its antidepressant properties. Following a 30-minute interval after extract administration, each mouse was placed in a water-filled chamber, maintaining a temperature between 25 °C and 27 °C. During the final four minutes of the test, the immobility time of each mouse was meticulously recorded. This immobility duration serves as a crucial indicator of behaviors associated with depression-like.
Sucrose preference test: This assay was employed to assess the antidepressant qualities of protein extracts sourced from eggs and beans in the context of chronic mild depression [1414Krishnan V, Nestler EJ. Animal models of depression: molecular perspectives. Curr Top Behav Neurosci. 2011;7:121-47. doi: 10.1007/7854_2010_108. PMID: 21225412; PMCID: PMC3270071.]. The sucrose preference test, a widely recognized behavioral evaluation method introduced by Willner [1616Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology. 2005;52(2):90-110. doi: 10.1159/000087097. Epub 2005 Jul 19. PMID: 16037678.], was implemented for this investigation. To execute the experiment, the protein extracts were administered “i.p” to the mice on a daily basis, encompassing doses ranging from 14 µg/Kg (b.w.) to 140 µg/Kg (b.w.). In order to streamline the testing procedure, the mice were organized into 16 distinct groups. Concurrently, control groups were subjected solely to a physiological solution (0.9% NaCl, pH = 7.4), while additional groups of mice were treated with sertraline, reference molecule. Subsequent to the extracts administration, the mice were individually housed in cages, each furnished with two separate bottles: one containing water and the other a 1% sucrose solution. The daily consumption of both water and the sucrose solution was methodically measured and recorded. To mitigate potential confusion, the positions of the two bottles were alternated daily. Calculation of sucrose preference was conducted using the following formula:
Statistical analysis
The results acquired were presented as the mean ± standard error of the mean (SEM). To contrast the means among various groups, the Anova test was utilized. A finding was deemed statistically significant if the p - value was below 0.05 (p < 0.05).
Table 1 displays the primary morphological and behavioral changes following the administration of undenatured protein extracts derived from beans at saturation levels of 40% and 80%, spanning a range of doses. Aside from the observed fur alterations and increased restlessness among the animals (Table 1), no further behavioral changes were detected throughout the 14-day duration of the study, even at the highest administered dose and saturation levels (5000 mg/Kg, b.w.; and 80% saturation).
Table 1: Morphological and behavioral changes resulting from the administration of undenatured bean protein extracts.
- Effects of undenatured egg protein extracts
Following the administration of undenaturedegg protein extracts at saturation levels of 40% and 80% across various dosages, noticeable changes in the appearance of fur and heightened restlessness in the animals were recorded (Table 2). Specifically, some mice displayed raised fur spikes on the seventh day subsequent to receiving a dosage of 5000 mg/Kg (b.w.) of the 40% protein extract. Similar alterations in appearance were observed in the animals on the twelfth day of the study when administered a dosage of 700 mg/Kg (b.w.) of the same extract. Nevertheless, no additional shifts in behavior were documented over the course of the 14-day study period, even in cases involving the most concentrated protein extract (80%) at the highest dosage (5000mg/Kg, b.w.) (Table 2).
Table 2: Morphological and behavioral changes of the animals following the administration of undenatured egg protein extracts.
Weight evolution
- Effects of undenaturedbean protein extracts
Table 3 presents the results pertaining to the weight changes of the animals following the administration of undenatured bean protein extracts at saturation of40% and 80%, across different dosages. Monitoring the weight modifications in the treated animals did not reveal any weight-related toxic effects when compared to the control group. Insignificant weight fluctuations, whether in terms of gain or loss, were observed throughout the two-week study period among all groups, including the control group. It is important to note that these weight variations are not associated with the bean extracts under investigation (Table 3).
Table 3: Average body weight of the animals after receiving undenaturedbean protein extracts administration.
- Effects of undenatured egg protein extracts
As shown in Table 4, the weight progression of the animals subjected to undenatured egg protein extracts showed no significant changes. When compared to the control group, there were no observed toxic effects related to the weight of these animals. This indicates that the administration of undenatured egg protein extracts did not result in any adverse weight-related outcomes.
Table 4: Animals' body weight following the administration of undenatured egg protein extracts.
Organ weights
- Effects of undenatured bean protein extracts
The results concerning the effects of undenatured bean protein extracts on the vital organs of the animals indicated no significant differences in the relative weights of these organs (Table 5). In comparison to the control group, undenatured bean protein extracts at saturation levels of 40% and 80% did not induced any toxic effects on the vital organs, even at the highest dose (5000 mg/Kg, b.w.) (Table 5).
Table 5: Average organ weights of the animals following the administration of undenatured bean protein extracts.
- Effects of undenatured egg protein extracts
Table 6 displays the organ weights of the animals after the administration of undenatured egg protein extracts at saturation levels of 40% and 80% at various doses. Notably, the egg protein extracts at both 40% and 80% saturation were exhibiting no adverse impacts on the vital organs. Even at the highest administered dose (5000 mg/Kg, b.w.), there were no noteworthy differences compared to the control group (Table 6).
Table 6: Organ weights of the animals after the administration of undenatured egg protein extracts.
Mortality and LD50
- Effects of undenaturedbean protein extracts
The information provided in Table 7 offers insight into the occurrence of mouse fatalities subsequent to the administration of undenatured bean protein extracts. An examination of the acute toxicity of these extracts revealed a concerning finding two deaths were noted within the initial week after administration of undenatured bean protein extract at saturation of 80%, specifically at the doses of 2000 mg/Kg (b.w.) and 5000 mg/Kg (b.w.) (Table 7). Conversely, contrasting results were observed in the case of the 40% protein extract, where no instances of mortality were recorded at any of the investigated doses (Table 7).
Table 7: Mouse mortality number after administration of undenatured bean protein extracts.
- Effects of undenatured egg protein extracts
Upon conducting an assessment of the acute toxicity of undenatured egg protein extracts, no instances of mortality were recorded subsequent to the administration of protein extracts at both 40% and 80% saturation levels, regardless of the doses investigated.
Antioxidant activity of protein extracts
DPPH free radical scavenging: The outcomes of the DPPH test were inconclusive, as this particular assay neither confirmed nor negated the antioxidant activity of denatured and undenatured bean and egg protein extracts. This uncertainty arose from the fact that proteins were precipitating within the solvent used, which was ethanol. This phenomenon was acting as a limiting step in the antioxidant reaction.
H2O2 free radical scavenging: The provided data in Table 8 illustrates the antioxidant capacities of undenatured and denatured protein extracts derived from beans and eggs at different saturation levels. Additionally, the results obtained indicated that denatured protein extracts derived from beans and eggs demonstrated strong antioxidant activity when compared to undenatured protein extracts at various saturation levels (Table 8). For the bean protein extracts, it is evident that undenatured extracts at a saturation level of 80% had a significantly lower IC50 value of 56 ± 0.002 µg/mL compared to the IC50 value of 78.9 ± 0.03 µg/mL for the same extracts at a saturation level of 40% (Table 8). This implied that undenatured bean protein extracts at 80% saturation were more potent antioxidants, as they required a lower concentration to inhibit oxidative damage. Similarly, denatured bean protein extracts also exhibited enhanced antioxidant potential at higher saturation. The IC50 value for denatured bean protein extracts was 47.1 ± 0.04 µg/mL at 80% saturation, whereas it was 63 ± 0.06 µg/mL at 40% saturation (Table 8). Turning to egg protein extracts, the trend was consistent. Undenatured extracts showed greater antioxidant activity at 80% saturation (IC50 = 66.4 ± 0.08 µg/mL) compared to 40% saturation (IC50 = 87 ± 0.015 µg/mL). Similarly, denatured egg protein extracts displayed improved antioxidant potential at 80% saturation (IC50 = 51 ± 0.034 µg/mL) compared to 40% saturation (IC50 = 69 ± 0.007 µg/mL) (Table 8).
Table 8: Antioxidant capacities of undenatured and denatured protein extracts derived from beans and eggs at different saturation levels.
Acute antidepressant effect of protein extracts
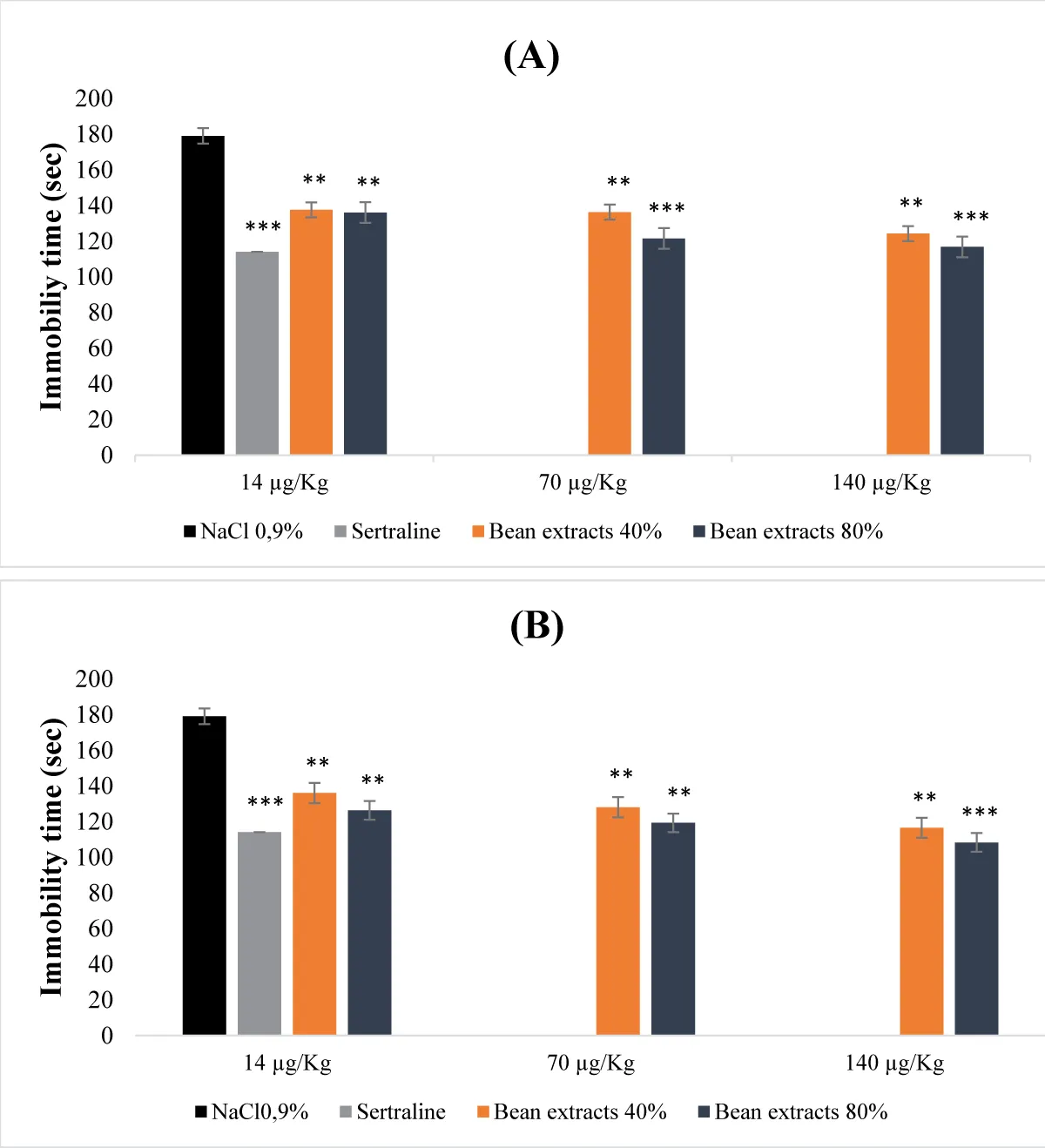
- Effect of bean protein extracts
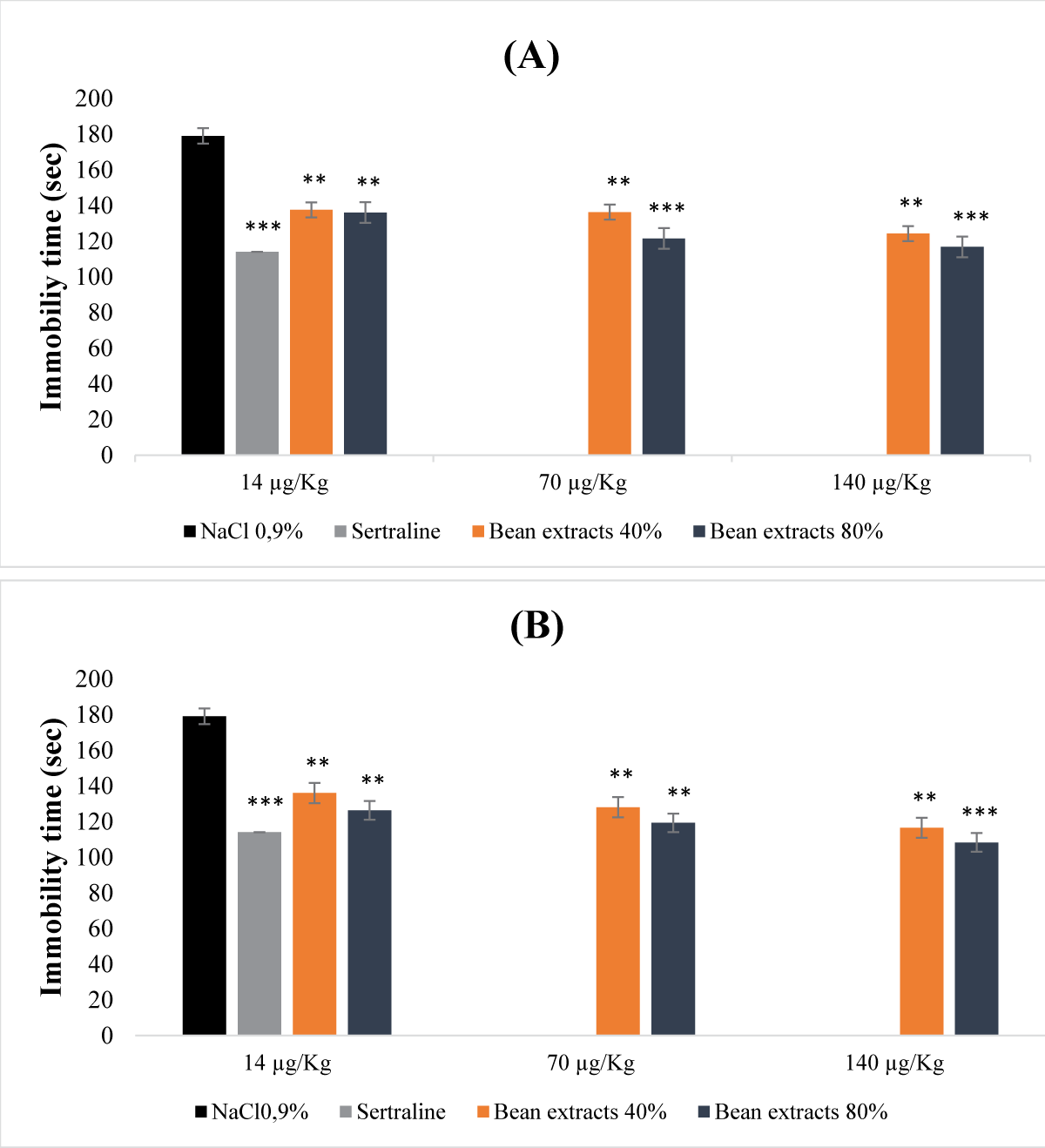
The obtained results were revealing that bean protein extracts were exhibiting a dose-dependent antidepressant activity (Figure 1). This activity was varying based on the type of extract used (Figures 1A,B). Undenatured 40% and 80% protein extracts were leading to a significant decrease in immobility time across all administered doses. This reduction was particularly prominent at the doses of 70 µg/Kg (b.w.) and 140 µg/Kg (b.w.) for the 80% protein extract (p <0.001). As for the denatured protein extracts, the reduction in immobility time was even more pronounced. In fact, as the dose increased, the immobility time decreased. Similar to the undenatured extracts, the reduction was notably significant for the 80% extract at the dose of 140 µg/Kg (b.w.) (Figure 1B). The antidepressant effect that was demonstrated by the undenatured 80% protein extract closely resembled that of the animals treated with sertraline (reference molecule). When compared to the control group, the effects of both denatured and undenatured bean protein extracts at saturation of 80% were more pronounced than those of sertraline (reference molecule) (Figure 1A,B).
Figure 1: Acute antidepressant effect of bean protein extracts. (A): Undenatured bean extracts; (B): Denatured bean extracts.
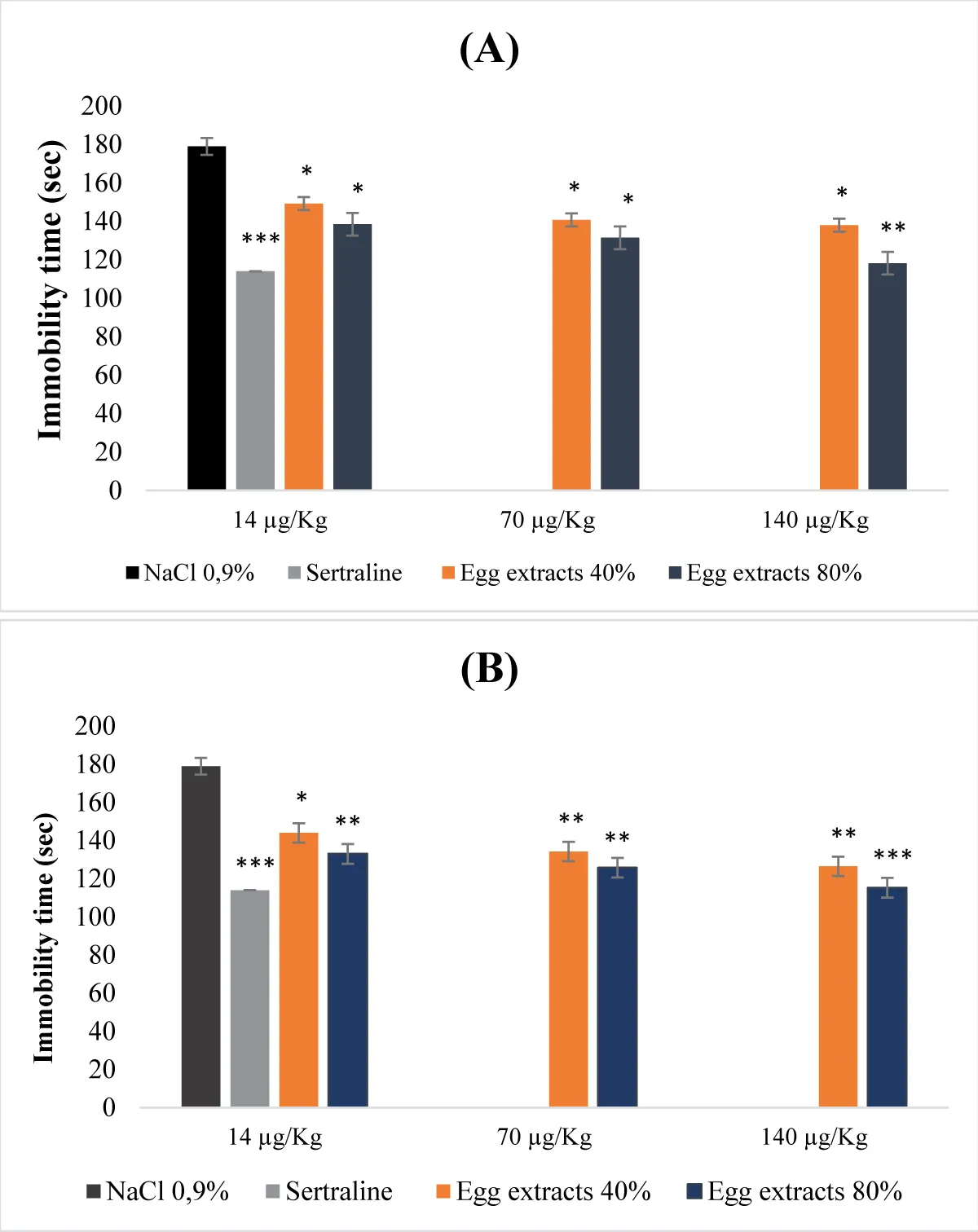
- Effect of egg protein extracts
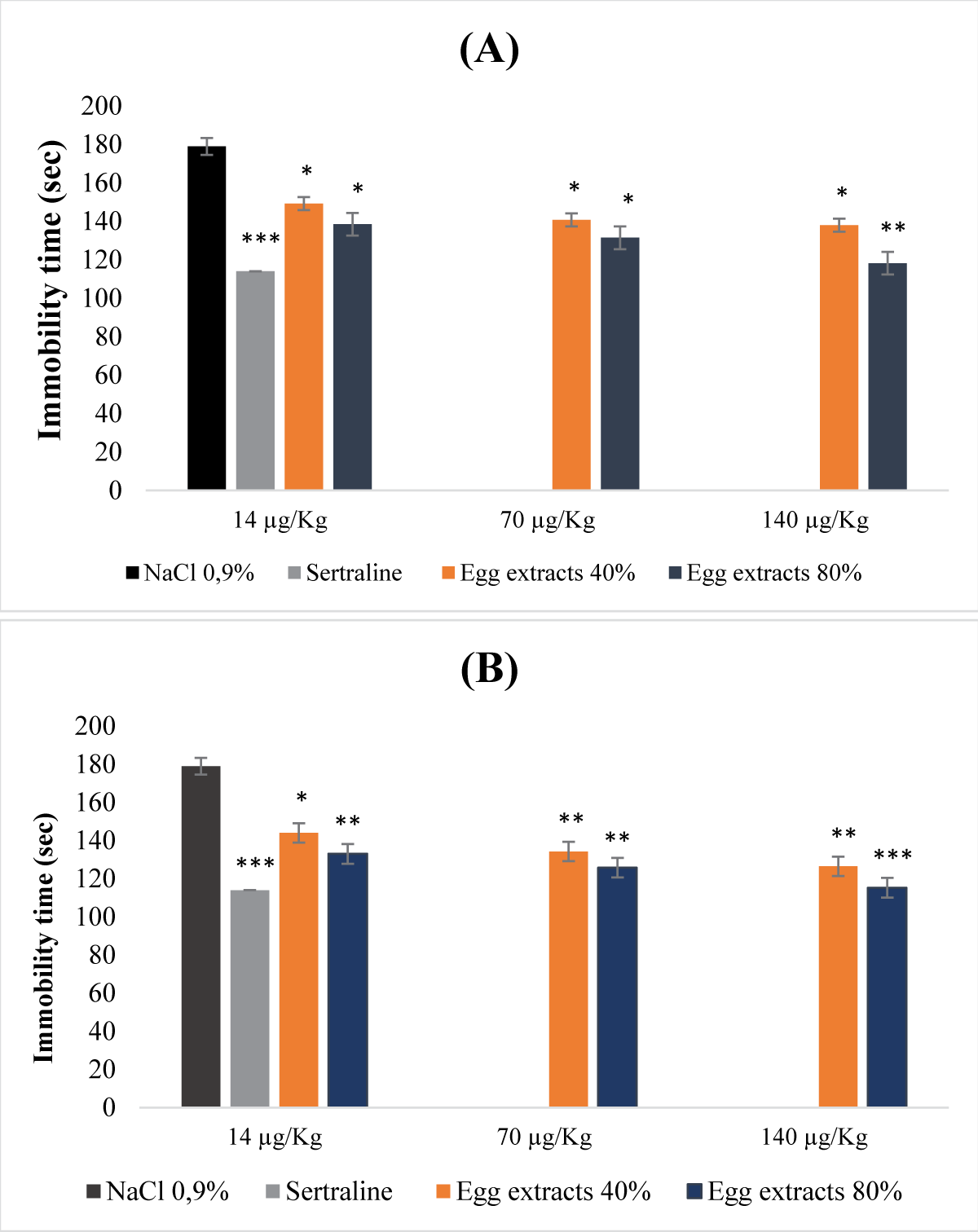
Figure 2 illustrates the acute antidepressant effect of egg protein extracts. The egg protein extracts at both 40% and 80% saturation levels exhibited a dose-dependent antidepressant effect. All extracts, whether denatured or not, displayed a reduction in immobility time inversely proportional to the studied extract doses (Figure 2A and B). This reduction was more pronounced when the extracts were denatured and the saturation percentage was 80% (Figure 2B). Indeed, treatment with the denatured 80% extract at the dose of 140μg/Kg (b.w.) resulted in a highly significant decrease in immobility time (p <0.001).
Figure 2: Acute antidepressant effect of egg protein extracts. (A): Undenatured bean extracts; (B): Denatured bean extracts.
Chronic antidepressant effect of protein extracts
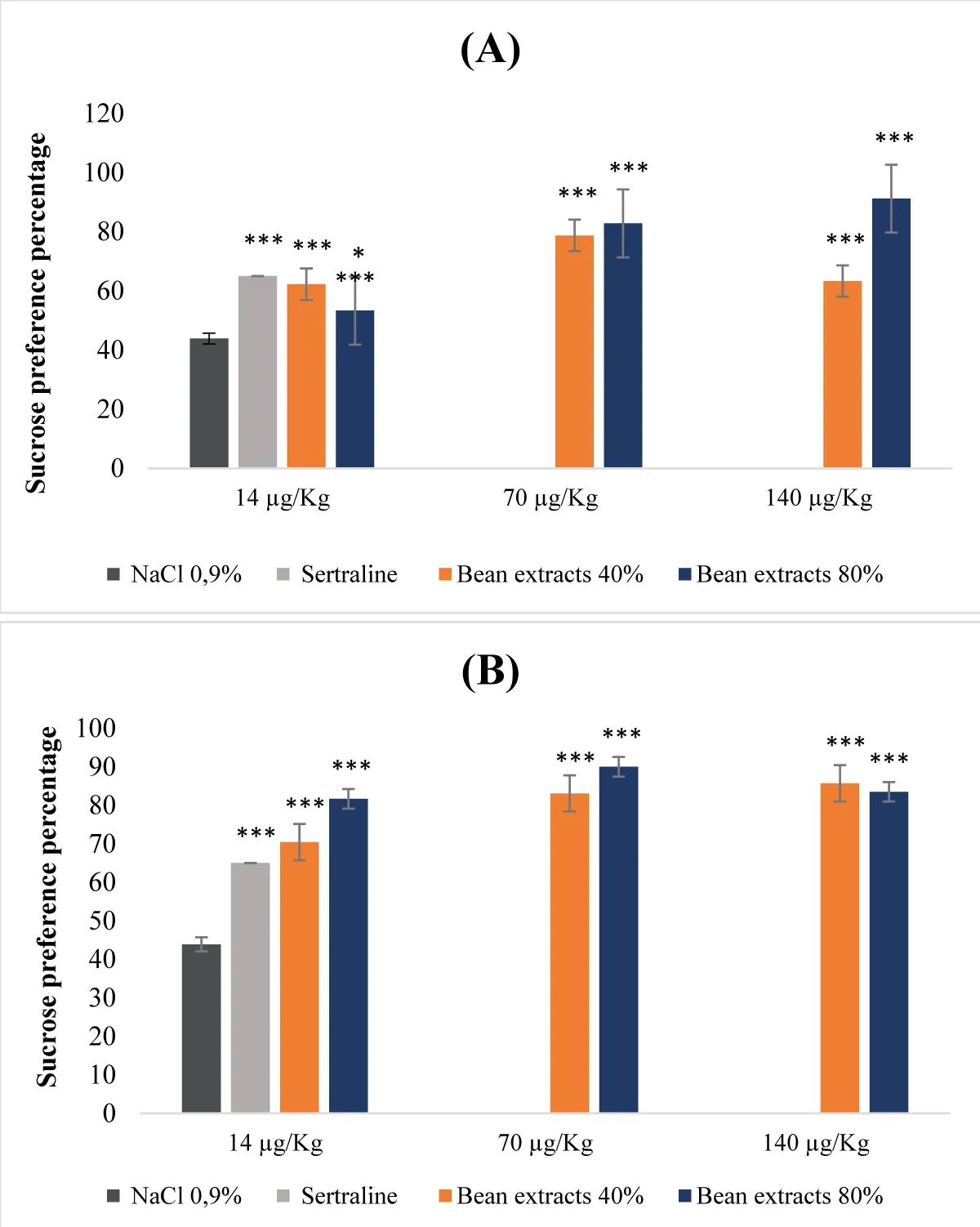
- Effect of bean protein extracts
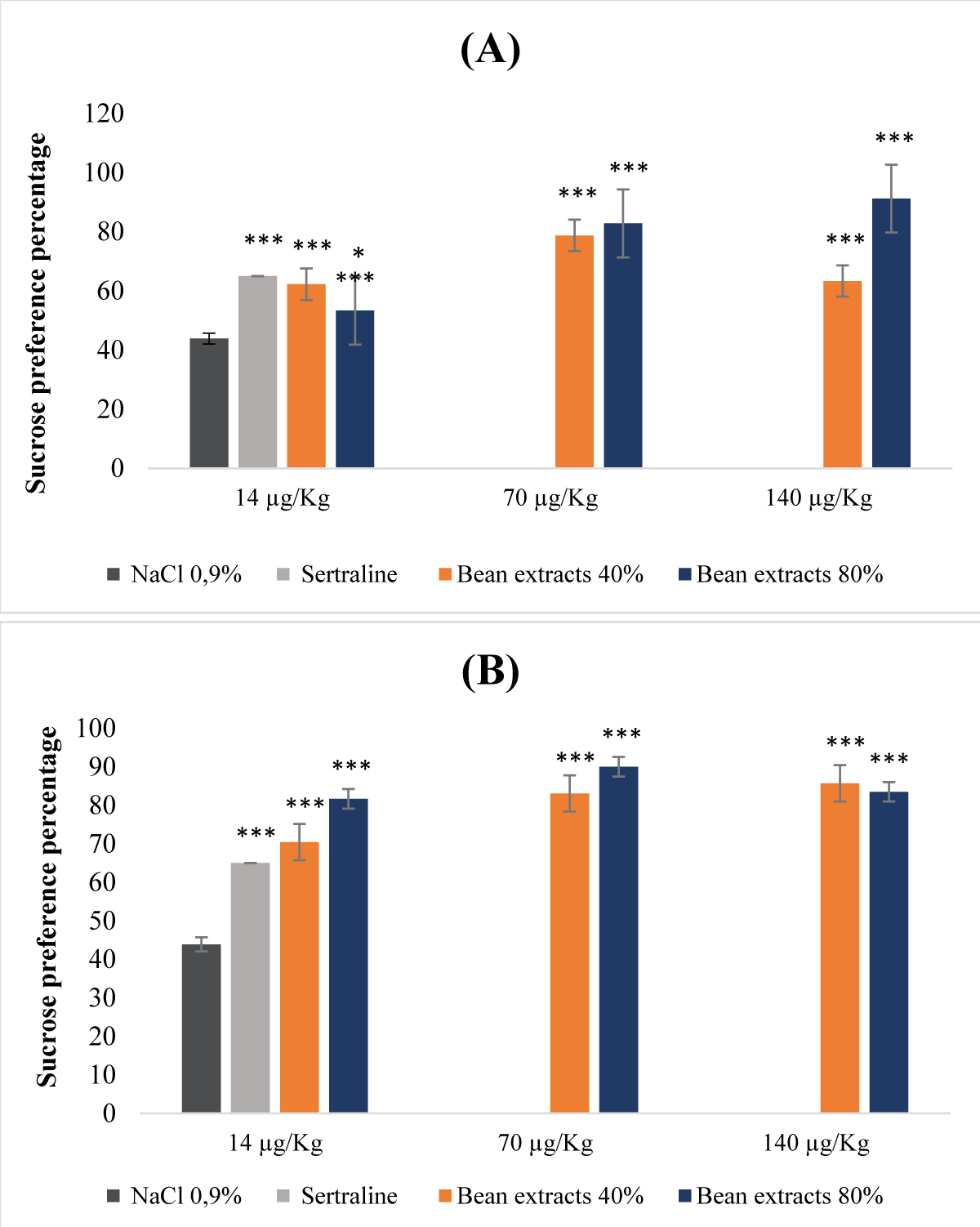
Interpreting the results obtained from the data presented in the Figure 3, protein extracts derived from beans at saturation levels of both 40% and 80%, regardless of whether they were denatured or not, displayed a dose-dependent antidepressant impact (Figure 3A,B). For the non-denatured protein extracts, the groups treated with the former exhibited a notably elevated sucrose preference rate (Figure 3A). This increase was particularly conspicuous in the cohort treated with the 80% saturation protein extract, administered at a dose of 140 µg/Kg (b.w.). Notably, the effect of this particular extract even out performed that of sertraline (Figure 3A). Simultaneously, the 40% saturation protein extract, administered at a dose of 70 µg/Kg (b.w.), demonstrated a more pronounced effect when contrasted with the reference molecule. In the context of denatured extracts, all tested doses, at both 40% and 80% saturation levels, revealed a significantly substantial sucrose preference percentage (p <0.001), surpassing even that observed in animals treated with sertraline (Figure 3B).
Figure 3: Antidepressant properties of protein extracts derived from beans. (A): Undenatured bean extracts; (B): Denatured bean extracts.
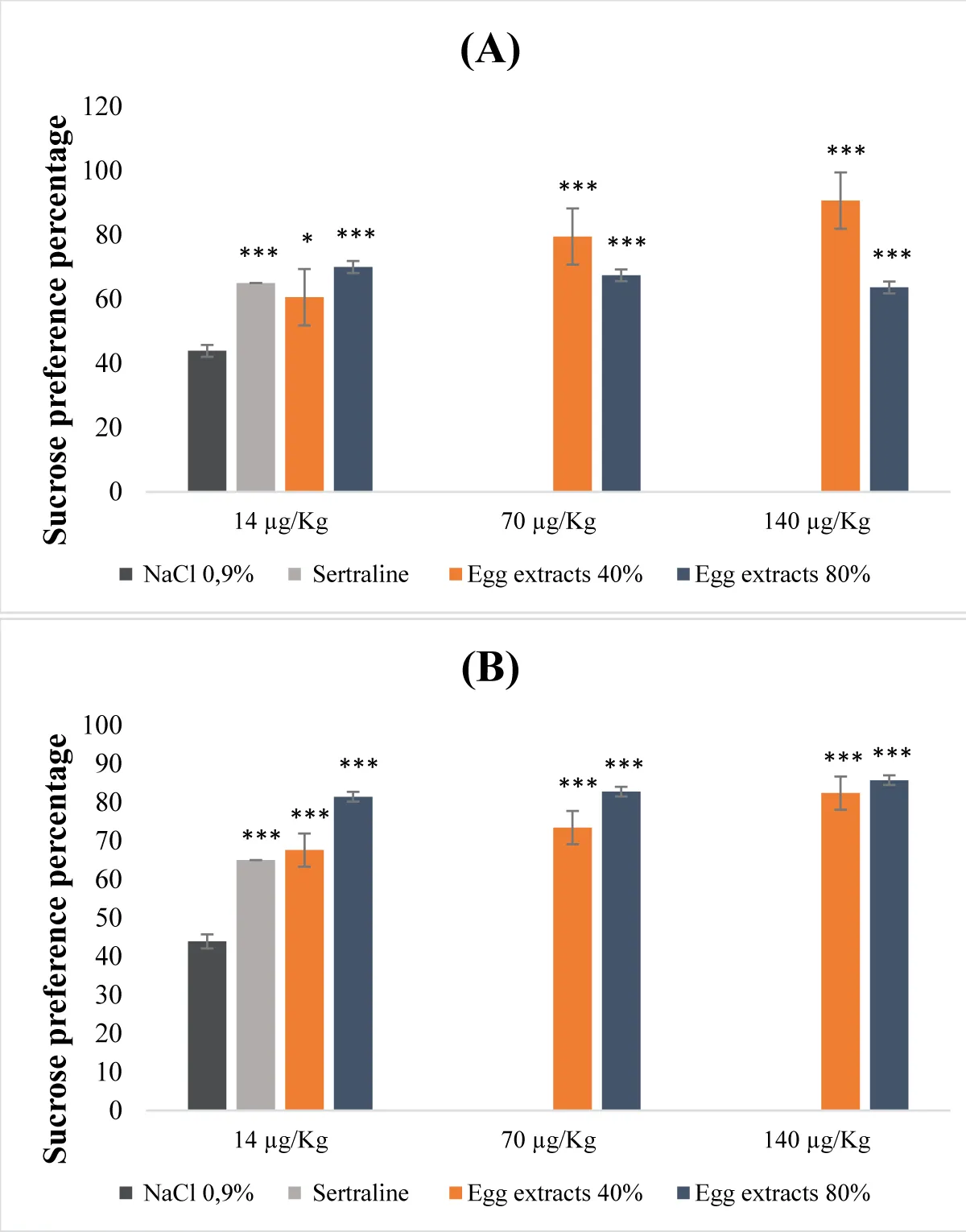
- Effect of egg protein extracts
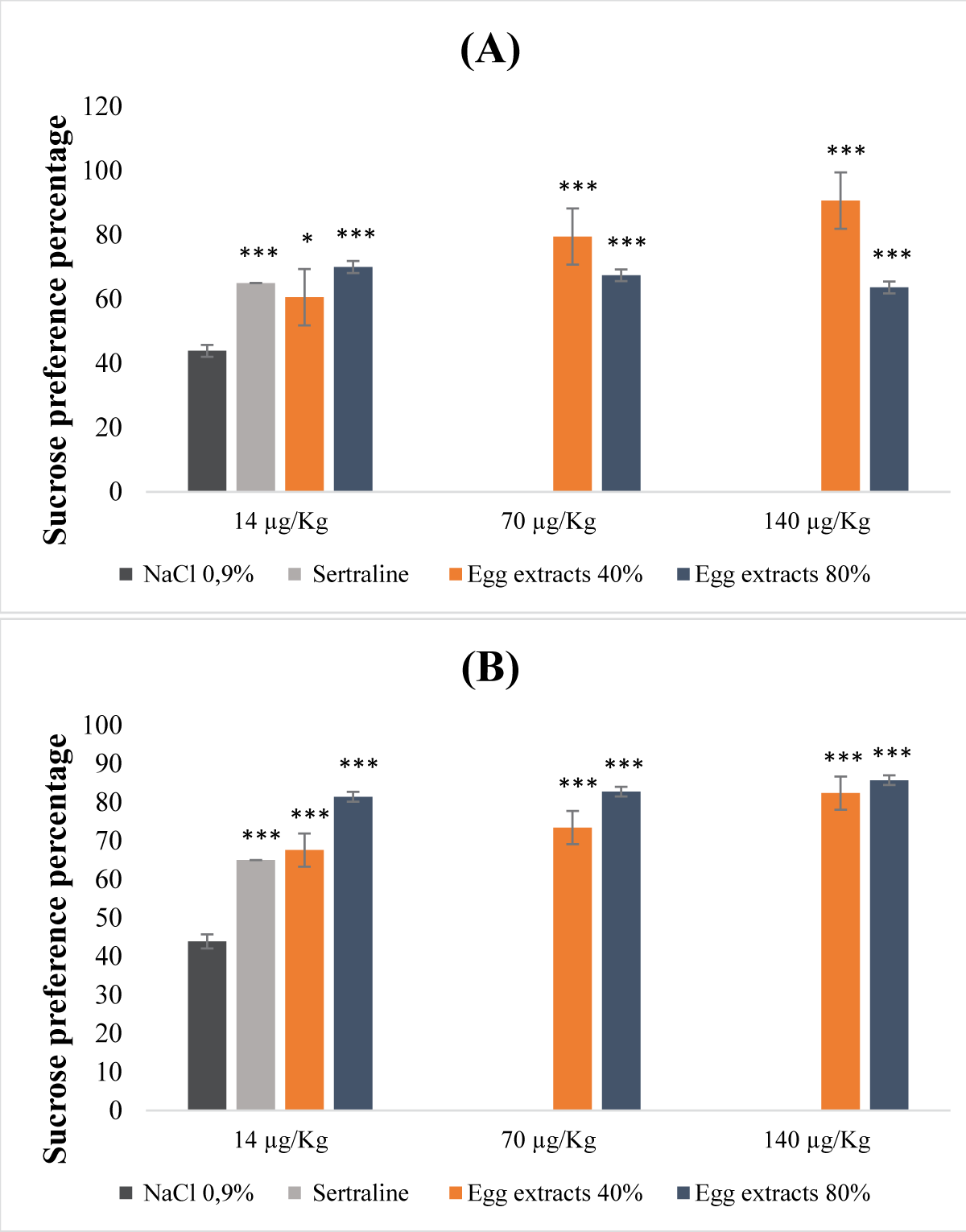
Egg protein extracts exhibited a dose-dependent response in their antidepressant activity (Figure 4). This potential to alleviate depression varied based on the types of extracts examined, including saturations of 40% and 80%, whether they were subject to denaturation or not (Figure 4A,B). In the case of denatured egg extracts with an 80% saturation level, the antidepressant effect was notably significant (p <0.001) across all administered dosage levels. Specifically, the sucrose preference percentages reached 71%, 83%, and 86% for doses of 14, 70, and 140 µg/Kg (b.w.), respectively (Figure 4B). Conversely, for the undenatured extracts, the 40% saturation extract displayed a more substantial antidepressant impact compared to the 80% saturation counterpart, particularly evident at doses of 70 and 140 µg/Kg (b.w.) (Figure 4A).
Figure 4: Antidepressant properties of protein extracts derived from egg (A): Undenatured bean extracts; (B): Denatured bean extracts.
This research highlights, for the first time, the pharmacognostic antidepressant potential present in nutrients sourced from both legume and animal origins proteins. To accomplish this, preclinical investigations were carried out using mice animal models. Moreover, antioxidant assessments were conducted to uncover the antioxidative capabilities of these nutrients.The selection of these nutrients was guided by multiple scientific criteria, which included; (i) The underlying pathophysiology of depression, and (ii) the enrichment of protein extracts with polypeptides, peptides, and amino acids that could serve as precursors to neurotransmitters implicated in depression.
For these reasons, we delved into exploring the pharmacognostic antidepressant attributes of non-denatured protein extracts sourced from beans and eggs at saturation levels of 40% and 80%, categorizing them as macronutrients. Additionally, we investigated denatured protein extracts from beans and eggs at saturation levels of 40% and 80% as micronutrients.
The evaluation of the antioxidant activity in bean and egg protein extracts, whether they were denatured or not, utilizing the DPPH test, did not yield conclusive evidence either confirming or refuting the anticipated impacts of the examined nutrients. This observation can be attributed to the precipitative influence of the ethanol employed in this test, as highlighted by Wessel and Flügge [1717Wessel D, Flügge UI. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal Biochem. 1984 Apr;138(1):141-3. doi: 10.1016/0003-2697(84)90782-6. PMID: 6731838.]. Ethanol induces protein precipitation by dehydrating them, thereby limiting their reactivity with DPPH in the reaction medium, subsequently impeding the accurate assessment of their antioxidant potential. In accordance with this assay, our findings are consistent with the research conducted by Fic, et al. and that of Kita, et al. [1818Fic E, Kedracka-Krok S, Jankowska U, Pirog A, Dziedzicka-Wasylewska M. Comparison of protein precipitation methods for various rat brain structures prior to proteomic analysis. Electrophoresis. 2010 Oct;31(21):3573-9. doi: 10.1002/elps.201000197. PMID: 20967768.,1919Kita Y, Arakawa T, Lin TY, Timasheff SN. Contribution of the surface free energy perturbation to protein-solvent interactions. Biochemistry. 1994 Dec 20;33(50):15178-89. doi: 10.1021/bi00254a029. PMID: 7999778.]. Concerning the H2O2 test, the analysis of bean and egg protein extracts at saturation levels of 40% and 80%, regardless of denaturation status, revealed a substantial antioxidant capacity. This noteworthy antioxidative potential is likely attributed to the presence of proteins and peptides rich in tryptophan (Trp), tyrosine (Tyr), methionine, and cysteine (Cys). Research by Torkova, et al. and Hernández-Ledesma, et al. [2020Torkova A, Koroleva O, Khrameeva E, Fedorova T, Tsentalovich M. Structure-Functional Study of Tyrosine and Methionine Dipeptides: An Approach to Antioxidant Activity Prediction. Int J Mol Sci. 2015 Oct 23;16(10):25353-76. doi: 10.3390/ijms161025353. PMID: 26512651; PMCID: PMC4632805.,2121Hernández-Ledesma B, Dávalos A, Bartolomé B, Amigo L. Preparation of antioxidant enzymatic hydrolysates from alpha-lactalbumin and beta-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J Agric Food Chem. 2005 Feb 9;53(3):588-93. doi: 10.1021/jf048626m. PMID: 15686406.] underscores the varying antioxidant abilities of amino acids Trp, Tyr, Met, and Cys, contingent upon their specific nature and whether they reside at the N- or C-terminal position. Furthermore, studies indicate that dipeptides featuring Tyr and Trp at the N-terminal exhibit more robust antioxidant activities compared to those positioned at the C-terminal end [2222Zheng L, Zhao Y, Dong H, Su G, Zhao M. Structure–activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. Journal of Functional Foods. 2016; 21:485‑]. Interestingly, various antioxidant dipeptides, such as Trp-Tyr and Met-Trp, have demonstrated intramolecular synergistic effects [2323Güngör N, Ozyürek M, Güçlü K, Cekiç SD, Apak R. Comparative evaluation of antioxidant capacities of thiol-based antioxidants measured by different in vitro methods. Talanta. 2011 Feb 15;83(5):1650-8. doi: 10.1016/j.talanta.2010.11.048. Epub 2010 Nov 30. PMID: 21238764.-2525Cano A, Alcaraz O, Arnao MB. Free radical-scavenging activity of indolic compounds in aqueous and ethanolic media. Anal Bioanal Chem. 2003 May;376(1):33-7. doi: 10.1007/s00216-003-1848-7. Epub 2003 Mar 29. PMID: 12734615.]. Utilizing this gathered data, it becomes apparent that hydrogen peroxide, a molecule produced during immune responses and metabolic processes [2626Lennicke C, Rahn J, Lichtenfels R, Wessjohann LA, Seliger B. Hydrogen peroxide - production, fate and role in redox signaling of tumor cells. Cell Commun Signal. 2015 Sep 14;13:39. doi: 10.1186/s12964-015-0118-6. PMID: 26369938; PMCID: PMC4570748.], assumes significant roles. While essential for various physiological functions, this molecule also exhibits neurotoxic properties by inhibiting neurons within the paraventricular nucleus of the hypothalamus through the activation of potassium channels [2727Pouokam E, Rehn M, Diener M. Effects of H2O2 at rat myenteric neurones in culture. Eur J Pharmacol. 2009 Aug 1;615(1-3):40-9. doi: 10.1016/j.ejphar.2009.04.066. Epub 2009 May 14. PMID: 19446543.,2828Dantzler HA, Matott MP, Martinez D, Kline DD. Hydrogen peroxide inhibits neurons in the paraventricular nucleus of the hypothalamus via potassium channel activation. Am J Physiol Regul Integr Comp Physiol. 2019 Jul 1;317(1):R121-R133. doi: 10.1152/ajpregu.00054.2019. Epub 2019 May 1. PMID: 31042419; PMCID: PMC6692749.]. Considering the demonstrated capacity of both denatured and non-denatured extracts from beans and eggs to counteract the effects of H2O2, a hypothesis arises suggesting the potential neuroprotective qualities of these extracts, particularly concerning the influence of hydrogen peroxide. This intriguing notion raises the possibility that these protein extracts might potentially contribute to the preservation of neural well-being, especially in the context of H2O2-induced neurotoxicity.
Upon completion of the acute toxicity study, it became apparent that when administered at doses of 2000 mg/Kg (b.w.) and 5000 mg/Kg (b.w.), the bean protein extract at an 80% saturation level exhibited toxicity. This harmful effect was confirmed by instances of mortality observed following the administration of these specific doses. In contrast, regardless of the dosage levels examined, egg protein extracts at 40% and 80% saturation did not display any indications of toxicity. As a result, the calculated LD50 values for the egg protein extracts and the bean protein extract at 40% saturation exceeded 5g/Kg (b.w.), indicating their lack of toxicity. However, the LD50 value for the bean protein extract at 80% saturation was determined to be 2.5 g/Kg (b.w.), indicating a relatively higher level of toxicity (OCED) [2929Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015 Mar 2;(97):52587. doi: 10.3791/52587. PMID: 25867960; PMCID: PMC4401172.]. Consequently, based on the established LD50 values, it can be concluded that the egg protein extracts at 40% and 80% saturation, as well as the bean protein extract at 40% saturation, can be classified as non-toxic substances [2929Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015 Mar 2;(97):52587. doi: 10.3791/52587. PMID: 25867960; PMCID: PMC4401172.]. Nevertheless, it is important to highlight that the bean protein extract at 80% saturation may be categorized as having a mild level of toxicity [2929Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015 Mar 2;(97):52587. doi: 10.3791/52587. PMID: 25867960; PMCID: PMC4401172.].
In the examination of the pharmacognostic qualities linked to the antidepressant attributes of the studied nutrients, a comprehensive analysis involved the utilization of two distinct depression tests. The primary objective of the initial test (FST) was to assess the effectiveness of these nutrients in managing instances of acute depression [1414Krishnan V, Nestler EJ. Animal models of depression: molecular perspectives. Curr Top Behav Neurosci. 2011;7:121-47. doi: 10.1007/7854_2010_108. PMID: 21225412; PMCID: PMC3270071.], while the second evaluation (sucrose preference test) delved into their potential in the treatment of chronic mild depression conditions [1414Krishnan V, Nestler EJ. Animal models of depression: molecular perspectives. Curr Top Behav Neurosci. 2011;7:121-47. doi: 10.1007/7854_2010_108. PMID: 21225412; PMCID: PMC3270071.]. These behavioral assessments, widely adopted in the investigation of antidepressant agents [], stood as pivotal benchmarks. Concentrating on acute depression, a substantial and noteworthy antidepressant effect emerged subsequent to the administration of denatured bean and egg protein extracts at a saturation level of 80%. This finding underscores the robust potential of denatured proteins, particularly those sourced from beans and eggs at an 80% saturation point, in addressing acute depressive states. This outcome harmonizes with previous research by Van Praag, Sandyk, Young, and Meyers [3131Sandyk R. L-tryptophan in neuropsychiatric disorders: a review. Int J Neurosci. 1992 Nov-Dec;67(1-4):127-44. doi: 10.3109/00207459208994781. PMID: 1305630.-3434Treadway MT, Zald DH. Reconsidering anhedonia in depression: lessons from translational neuroscience. Neurosci Biobehav Rev. 2011 Jan;35(3):537-55. doi: 10.1016/j.neubiorev.2010.06.006. Epub 2010 Jul 11. PMID: 20603146; PMCID: PMC3005986.], who unearthed promising outcomes tied to tyrosine and tryptophan within the domain of depressive therapy. The engagement of peptides and amino acids originating from these denatured proteins is postulated as an influential mechanism. This notion revolves around the concept that these biologically active components may function by counterbalancing deficiencies in neurotransmitter precursors (dopamine, serotonin, etc.). Through the infusion of amino acids inherent in denatured protein compounds, like the neurotransmitter precursors tyrosine and tryptophan both fundamental constituents for an array of neurotransmitters encompassing dopamine and serotonina plausible avenue for mitigating neurotransmitter imbalances is suggested. Additionally, this intricate process might extend to the enhancement of neuronal membrane flexibility, coupled with the amplification of neurotransmitter receptors integral to the intricate tapestry of depression. In the domain of chronic depression, a significant aspect comes to the forefront: denatured bean and egg protein extracts, with a saturation of 80%, demonstrated a remarkable ability to alleviate anhedonia, a primary manifestation of depressive conditions [3535Schalla MA, Kühne SG, Friedrich T, Hanel V, Kobelt P, Goebel-Stengel M, Rose M, Stengel A. Sucrose Preference and Novelty-Induced Hypophagia Tests in Rats using an Automated Food Intake Monitoring System. J Vis Exp. 2020 May 8;(159). doi: 10.3791/60953. PMID: 32449711.]. This observation draws a parallel between these protein extracts and conventional antidepressants, both of which exhibit the potential to counter anhedonia by fostering positive cognitions and intensifying the experience of pleasure [3636Liu Y, Zhao J, Guo W. Emotional Roles of Mono-Aminergic Neurotransmitters in Major Depressive Disorder and Anxiety Disorders. Front Psychol. 2018 Nov 21;9:2201. doi: 10.3389/fpsyg.2018.02201. PMID: 30524332; PMCID: PMC6262356.]. These noteworthy findings amplify the inherent antidepressant potential present in the scrutinized micronutrients when utilized as protein-based agents, effectively recalibrating the intricate interplay of neurotransmitter anomalies at the core of depression’s biological underpinnings. The earlier explained rationales retain their relevance within this context, further fortified by the concept that denatured protein extracts at a saturation of 80% function as pivotal foundational components for neurotransmitter synthesis, particularly concerning dopamine and serotonin. A wealth of investigations has illuminated the role played by nerve fibers originating from dopaminergic and serotonergic neurons, transmitting vital nerve impulses to regions such as the cerebral cortex and hippocampus. This intricate orchestration culminates in the release of neurotransmitters dopamine (DA) and serotonin (5-HT), pivotal in overseeing a spectrum of functions encompassing emotional modulation, motivation, memory, and a range of pleasurable experiences [3636Liu Y, Zhao J, Guo W. Emotional Roles of Mono-Aminergic Neurotransmitters in Major Depressive Disorder and Anxiety Disorders. Front Psychol. 2018 Nov 21;9:2201. doi: 10.3389/fpsyg.2018.02201. PMID: 30524332; PMCID: PMC6262356.].
This study highlights the antidepressant potential of undenatured bean and egg protein extracts as macronutrients, and denatured bean and egg protein extracts as micronutrients. Thus, these macronutrients and micronutrients may exert a neuroprotective effect due to their antioxidant power, neutralizing the impact of H2O2. Regarding toxicity, egg protein extracts were non-toxic, whereas bean protein extracts were mildly toxic. For these reasons, bean and egg protein extracts could constitute an inexhaustible source of new, effective, and harmless antidepressant molecules.
I would like to express my heartfelt gratitude to the all co-authors for their invaluable contributions and support throughout this endeavor. Your dedication and expertise have been instrumental in the successful completion of this work, and I am truly appreciative of your efforts. Thank you for your unwavering commitment and collaboration.
Smith K. The intricate relationship between diet, mental health, and well-being. Frontiers in Human Neuroscience. 2019; 13(33).
Sarris J, Logan AC, Akbaraly TN, Amminger GP, Balanzá-Martínez V, Freeman MP, Hibbeln J, Matsuoka Y, Mischoulon D, Mizoue T, Nanri A, Nishi D, Ramsey D, Rucklidge JJ, Sanchez-Villegas A, Scholey A, Su KP, Jacka FN; International Society for Nutritional Psychiatry Research. Nutritional medicine as mainstream in psychiatry. Lancet Psychiatry. 2015 Mar;2(3):271-4. doi: 10.1016/S2215-0366(14)00051-0. Epub 2015 Feb 25. PMID: 26359904.
Gómez-Pinilla F. Brain foods: the effects of nutrients on brain function. Nat Rev Neurosci. 2008 Jul;9(7):568-78. doi: 10.1038/nrn2421. PMID: 18568016; PMCID: PMC2805706.
Belmaker RH, Agam G. Major depressive disorder. N Engl J Med. 2008 Jan 3;358(1):55-68. doi: 10.1056/NEJMra073096. PMID: 18172175.
Jacka FN. Nutritional Psychiatry: Where to Next? EBioMedicine. 2017 Mar;17:24-29. doi: 10.1016/j.ebiom.2017.02.020. Epub 2017 Feb 21. PMID: 28242200; PMCID: PMC5360575.
Lieberman MD, Gaunt R, Gilbert DT, Trope Y. Reflexion and reflection: A social cognitive neuroscience approach to attributional inference. Advances in experimental social psychology. 2002; 34:199‑
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell. 4th Garland Science; 2002.
Fernstrom JD. Large neutral amino acids: dietary effects on brain neurochemistry and function. Amino Acids. 2013 Sep;45(3):419-30. doi: 10.1007/s00726-012-1330-y. Epub 2012 Jun 8. PMID: 22677921.
Fernstrom JD, Fernstrom MH. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J Nutr. 2007 Jun;137(6 Suppl 1):1539S-1547S; discussion 1548S. doi: 10.1093/jn/137.6.1539S. PMID: 17513421.
Boye JI, Aksay S, Roufik S, Ribereau S, Mondor M, Farnworth E, Rajamohamed SH. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Research International. 2010; 43:537‑
Reduan FH, Shaari RM, Sayuti NSA, Mustapha NM, Abu Bakar MZ, Sithambaram S, Hamzah H. Acute and subacute dermal toxicity of ethanolic extract of Melastoma malabathricumleaves in Sprague-Dawley rats. Toxicol Res. 2020 Mar 26;36(3):203-210. doi: 10.1007/s43188-019-00013-5. PMID: 32685424; PMCID: PMC7352010.
Qamar F,Naveed S, Faizi S, Sana A. Formulation and Evaluation of Natural Antioxidant Cream of Ocimum basilicum. Latin American Journal of Pharmacy. 2021; 40:2293‑
Porsolt RD, Le Pichon M, Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977 Apr 21;266(5604):730-2. doi: 10.1038/266730a0. PMID: 559941.
Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology. 2005;52(2):90-110. doi: 10.1159/000087097. Epub 2005 Jul 19. PMID: 16037678.
Wessel D, Flügge UI. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal Biochem. 1984 Apr;138(1):141-3. doi: 10.1016/0003-2697(84)90782-6. PMID: 6731838.
Fic E, Kedracka-Krok S, Jankowska U, Pirog A, Dziedzicka-Wasylewska M. Comparison of protein precipitation methods for various rat brain structures prior to proteomic analysis. Electrophoresis. 2010 Oct;31(21):3573-9. doi: 10.1002/elps.201000197. PMID: 20967768.
Kita Y, Arakawa T, Lin TY, Timasheff SN. Contribution of the surface free energy perturbation to protein-solvent interactions. Biochemistry. 1994 Dec 20;33(50):15178-89. doi: 10.1021/bi00254a029. PMID: 7999778.
Torkova A, Koroleva O, Khrameeva E, Fedorova T, Tsentalovich M. Structure-Functional Study of Tyrosine and Methionine Dipeptides: An Approach to Antioxidant Activity Prediction. Int J Mol Sci. 2015 Oct 23;16(10):25353-76. doi: 10.3390/ijms161025353. PMID: 26512651; PMCID: PMC4632805.
Hernández-Ledesma B, Dávalos A, Bartolomé B, Amigo L. Preparation of antioxidant enzymatic hydrolysates from alpha-lactalbumin and beta-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J Agric Food Chem. 2005 Feb 9;53(3):588-93. doi: 10.1021/jf048626m. PMID: 15686406.
Zheng L, Zhao Y, Dong H, Su G, Zhao M. Structure–activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. Journal of Functional Foods. 2016; 21:485‑
Güngör N, Ozyürek M, Güçlü K, Cekiç SD, Apak R. Comparative evaluation of antioxidant capacities of thiol-based antioxidants measured by different in vitro methods. Talanta. 2011 Feb 15;83(5):1650-8. doi: 10.1016/j.talanta.2010.11.048. Epub 2010 Nov 30. PMID: 21238764.
van Overveld FW, Haenen GR, Rhemrev J, Vermeiden JP, Bast A. Tyrosine as important contributor to the antioxidant capacity of seminal plasma. Chem Biol Interact. 2000 Jul 3;127(2):151-61. doi: 10.1016/s0009-2797(00)00179-4. PMID: 10936230.
Cano A, Alcaraz O, Arnao MB. Free radical-scavenging activity of indolic compounds in aqueous and ethanolic media. Anal Bioanal Chem. 2003 May;376(1):33-7. doi: 10.1007/s00216-003-1848-7. Epub 2003 Mar 29. PMID: 12734615.
Lennicke C, Rahn J, Lichtenfels R, Wessjohann LA, Seliger B. Hydrogen peroxide - production, fate and role in redox signaling of tumor cells. Cell Commun Signal. 2015 Sep 14;13:39. doi: 10.1186/s12964-015-0118-6. PMID: 26369938; PMCID: PMC4570748.
Pouokam E, Rehn M, Diener M. Effects of H2O2 at rat myenteric neurones in culture. Eur J Pharmacol. 2009 Aug 1;615(1-3):40-9. doi: 10.1016/j.ejphar.2009.04.066. Epub 2009 May 14. PMID: 19446543.
Dantzler HA, Matott MP, Martinez D, Kline DD. Hydrogen peroxide inhibits neurons in the paraventricular nucleus of the hypothalamus via potassium channel activation. Am J Physiol Regul Integr Comp Physiol. 2019 Jul 1;317(1):R121-R133. doi: 10.1152/ajpregu.00054.2019. Epub 2019 May 1. PMID: 31042419; PMCID: PMC6692749.
Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015 Mar 2;(97):52587. doi: 10.3791/52587. PMID: 25867960; PMCID: PMC4401172.
van Praag HM. In search of the mode of action of antidepressants. 5-HTP/tyrosine mixtures in depressions. Neuropharmacology. 1983 Mar;22(3 Spec No):433-40. doi: 10.1016/0028-3908(83)90193-4. PMID: 6304561.
Sandyk R. L-tryptophan in neuropsychiatric disorders: a review. Int J Neurosci. 1992 Nov-Dec;67(1-4):127-44. doi: 10.3109/00207459208994781. PMID: 1305630.
Young SN. Behavioral effects of dietary neurotransmitter precursors: basic and clinical aspects. Neurosci Biobehav Rev. 1996 Summer;20(2):313-23. doi: 10.1016/0149-7634(95)00022-4. PMID: 8811719.
Meyers S. Use of neurotransmitter precursors for treatment of depression. Altern Med Rev. 2000 Feb;5(1):64-71. PMID: 10696120.
Schalla MA, Kühne SG, Friedrich T, Hanel V, Kobelt P, Goebel-Stengel M, Rose M, Stengel A. Sucrose Preference and Novelty-Induced Hypophagia Tests in Rats using an Automated Food Intake Monitoring System. J Vis Exp. 2020 May 8;(159). doi: 10.3791/60953. PMID: 32449711.
Liu Y, Zhao J, Guo W. Emotional Roles of Mono-Aminergic Neurotransmitters in Major Depressive Disorder and Anxiety Disorders. Front Psychol. 2018 Nov 21;9:2201. doi: 10.3389/fpsyg.2018.02201. PMID: 30524332; PMCID: PMC6262356.
El Youbi AE, Boudaia O, Sekkout Z, El Amrani N, Radallah D. The Antioxidant and Antidepressant Properties of Dietary Proteins Derived from Egg and Bean Extracts and Their Acute Toxicity: A Journey from Nutrition to Pharmacognosy. IgMin Res. Nov 16, 2023; 1(1): 032-042. IgMin ID: igmin114; DOI: 10.61927/igmin114; Available at: www.igminresearch.com/articles/pdf/igmin114.pdf
Address Correspondence: Amal El Hamsas El Youbi, Department of Biology, Faculty of Sciences Ben M’Sik, University Hassan II, Casablanca, Morocco, Email: [email protected]
How to cite this article: El Youbi AE, Boudaia O, Sekkout Z, El Amrani N, Radallah D. The Antioxidant and Antidepressant Properties of Dietary Proteins Derived from Egg and Bean Extracts and Their Acute Toxicity: A Journey from Nutrition to Pharmacognosy. IgMin Res. Nov 16, 2023; 1(1): 032-042. IgMin ID: igmin114; DOI: 10.61927/igmin114; Available at: www.igminresearch.com/articles/pdf/igmin114.pdf
Figure 1: Acute antidepressant effect of bean protein extrac...
Figure 2: Acute antidepressant effect of egg protein extract...
Figure 3: Antidepressant properties of protein extracts deri...
Figure 4: Antidepressant properties of protein extracts deri...
Table 1: Morphological and behavioral changes resulting fro...
Table 2: Morphological and behavioral changes of the animal...
Table 3: Average body weight of the animals after receiving...
Table 4: Animals' body weight following the administration ...
Table 5: Average organ weights of the animals following the...
Table 6: Organ weights of the animals after the administrat...
Table 7: Mouse mortality number after administration of und...
Table 8: Antioxidant capacities of undenatured and denature...
Smith K. The intricate relationship between diet, mental health, and well-being. Frontiers in Human Neuroscience. 2019; 13(33).
Sarris J, Logan AC, Akbaraly TN, Amminger GP, Balanzá-Martínez V, Freeman MP, Hibbeln J, Matsuoka Y, Mischoulon D, Mizoue T, Nanri A, Nishi D, Ramsey D, Rucklidge JJ, Sanchez-Villegas A, Scholey A, Su KP, Jacka FN; International Society for Nutritional Psychiatry Research. Nutritional medicine as mainstream in psychiatry. Lancet Psychiatry. 2015 Mar;2(3):271-4. doi: 10.1016/S2215-0366(14)00051-0. Epub 2015 Feb 25. PMID: 26359904.
Gómez-Pinilla F. Brain foods: the effects of nutrients on brain function. Nat Rev Neurosci. 2008 Jul;9(7):568-78. doi: 10.1038/nrn2421. PMID: 18568016; PMCID: PMC2805706.
Belmaker RH, Agam G. Major depressive disorder. N Engl J Med. 2008 Jan 3;358(1):55-68. doi: 10.1056/NEJMra073096. PMID: 18172175.
Jacka FN. Nutritional Psychiatry: Where to Next? EBioMedicine. 2017 Mar;17:24-29. doi: 10.1016/j.ebiom.2017.02.020. Epub 2017 Feb 21. PMID: 28242200; PMCID: PMC5360575.
Lieberman MD, Gaunt R, Gilbert DT, Trope Y. Reflexion and reflection: A social cognitive neuroscience approach to attributional inference. Advances in experimental social psychology. 2002; 34:199‑
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell. 4th Garland Science; 2002.
Fernstrom JD. Large neutral amino acids: dietary effects on brain neurochemistry and function. Amino Acids. 2013 Sep;45(3):419-30. doi: 10.1007/s00726-012-1330-y. Epub 2012 Jun 8. PMID: 22677921.
Fernstrom JD, Fernstrom MH. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J Nutr. 2007 Jun;137(6 Suppl 1):1539S-1547S; discussion 1548S. doi: 10.1093/jn/137.6.1539S. PMID: 17513421.
Boye JI, Aksay S, Roufik S, Ribereau S, Mondor M, Farnworth E, Rajamohamed SH. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Research International. 2010; 43:537‑
Reduan FH, Shaari RM, Sayuti NSA, Mustapha NM, Abu Bakar MZ, Sithambaram S, Hamzah H. Acute and subacute dermal toxicity of ethanolic extract of Melastoma malabathricumleaves in Sprague-Dawley rats. Toxicol Res. 2020 Mar 26;36(3):203-210. doi: 10.1007/s43188-019-00013-5. PMID: 32685424; PMCID: PMC7352010.
Qamar F,Naveed S, Faizi S, Sana A. Formulation and Evaluation of Natural Antioxidant Cream of Ocimum basilicum. Latin American Journal of Pharmacy. 2021; 40:2293‑
Porsolt RD, Le Pichon M, Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977 Apr 21;266(5604):730-2. doi: 10.1038/266730a0. PMID: 559941.
Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology. 2005;52(2):90-110. doi: 10.1159/000087097. Epub 2005 Jul 19. PMID: 16037678.
Wessel D, Flügge UI. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal Biochem. 1984 Apr;138(1):141-3. doi: 10.1016/0003-2697(84)90782-6. PMID: 6731838.
Fic E, Kedracka-Krok S, Jankowska U, Pirog A, Dziedzicka-Wasylewska M. Comparison of protein precipitation methods for various rat brain structures prior to proteomic analysis. Electrophoresis. 2010 Oct;31(21):3573-9. doi: 10.1002/elps.201000197. PMID: 20967768.
Kita Y, Arakawa T, Lin TY, Timasheff SN. Contribution of the surface free energy perturbation to protein-solvent interactions. Biochemistry. 1994 Dec 20;33(50):15178-89. doi: 10.1021/bi00254a029. PMID: 7999778.
Torkova A, Koroleva O, Khrameeva E, Fedorova T, Tsentalovich M. Structure-Functional Study of Tyrosine and Methionine Dipeptides: An Approach to Antioxidant Activity Prediction. Int J Mol Sci. 2015 Oct 23;16(10):25353-76. doi: 10.3390/ijms161025353. PMID: 26512651; PMCID: PMC4632805.
Hernández-Ledesma B, Dávalos A, Bartolomé B, Amigo L. Preparation of antioxidant enzymatic hydrolysates from alpha-lactalbumin and beta-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J Agric Food Chem. 2005 Feb 9;53(3):588-93. doi: 10.1021/jf048626m. PMID: 15686406.
Zheng L, Zhao Y, Dong H, Su G, Zhao M. Structure–activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. Journal of Functional Foods. 2016; 21:485‑
Güngör N, Ozyürek M, Güçlü K, Cekiç SD, Apak R. Comparative evaluation of antioxidant capacities of thiol-based antioxidants measured by different in vitro methods. Talanta. 2011 Feb 15;83(5):1650-8. doi: 10.1016/j.talanta.2010.11.048. Epub 2010 Nov 30. PMID: 21238764.
van Overveld FW, Haenen GR, Rhemrev J, Vermeiden JP, Bast A. Tyrosine as important contributor to the antioxidant capacity of seminal plasma. Chem Biol Interact. 2000 Jul 3;127(2):151-61. doi: 10.1016/s0009-2797(00)00179-4. PMID: 10936230.
Cano A, Alcaraz O, Arnao MB. Free radical-scavenging activity of indolic compounds in aqueous and ethanolic media. Anal Bioanal Chem. 2003 May;376(1):33-7. doi: 10.1007/s00216-003-1848-7. Epub 2003 Mar 29. PMID: 12734615.
Lennicke C, Rahn J, Lichtenfels R, Wessjohann LA, Seliger B. Hydrogen peroxide - production, fate and role in redox signaling of tumor cells. Cell Commun Signal. 2015 Sep 14;13:39. doi: 10.1186/s12964-015-0118-6. PMID: 26369938; PMCID: PMC4570748.
Pouokam E, Rehn M, Diener M. Effects of H2O2 at rat myenteric neurones in culture. Eur J Pharmacol. 2009 Aug 1;615(1-3):40-9. doi: 10.1016/j.ejphar.2009.04.066. Epub 2009 May 14. PMID: 19446543.
Dantzler HA, Matott MP, Martinez D, Kline DD. Hydrogen peroxide inhibits neurons in the paraventricular nucleus of the hypothalamus via potassium channel activation. Am J Physiol Regul Integr Comp Physiol. 2019 Jul 1;317(1):R121-R133. doi: 10.1152/ajpregu.00054.2019. Epub 2019 May 1. PMID: 31042419; PMCID: PMC6692749.
Yankelevitch-Yahav R, Franko M, Huly A, Doron R. The forced swim test as a model of depressive-like behavior. J Vis Exp. 2015 Mar 2;(97):52587. doi: 10.3791/52587. PMID: 25867960; PMCID: PMC4401172.
van Praag HM. In search of the mode of action of antidepressants. 5-HTP/tyrosine mixtures in depressions. Neuropharmacology. 1983 Mar;22(3 Spec No):433-40. doi: 10.1016/0028-3908(83)90193-4. PMID: 6304561.
Sandyk R. L-tryptophan in neuropsychiatric disorders: a review. Int J Neurosci. 1992 Nov-Dec;67(1-4):127-44. doi: 10.3109/00207459208994781. PMID: 1305630.
Young SN. Behavioral effects of dietary neurotransmitter precursors: basic and clinical aspects. Neurosci Biobehav Rev. 1996 Summer;20(2):313-23. doi: 10.1016/0149-7634(95)00022-4. PMID: 8811719.
Meyers S. Use of neurotransmitter precursors for treatment of depression. Altern Med Rev. 2000 Feb;5(1):64-71. PMID: 10696120.
Schalla MA, Kühne SG, Friedrich T, Hanel V, Kobelt P, Goebel-Stengel M, Rose M, Stengel A. Sucrose Preference and Novelty-Induced Hypophagia Tests in Rats using an Automated Food Intake Monitoring System. J Vis Exp. 2020 May 8;(159). doi: 10.3791/60953. PMID: 32449711.
Liu Y, Zhao J, Guo W. Emotional Roles of Mono-Aminergic Neurotransmitters in Major Depressive Disorder and Anxiety Disorders. Front Psychol. 2018 Nov 21;9:2201. doi: 10.3389/fpsyg.2018.02201. PMID: 30524332; PMCID: PMC6262356.
The Antioxidant and Antidepressant Properties of Dietary Proteins Derived from Egg and Bean Extracts and Their Acute Toxicity: A Journey from Nutrition to Pharmacognosy



 Scan and get link
Scan and get link